
Respuestas del Sistema Pituitario - Adrenal - Gonadal al Sobreentrenamiento Provocado por el Entrenamiento de la Fuerza de Alta Intensidad
Leigh Ramsey1, William J Kraemer2 y A. C Fry1
1Human Performance Laboratories, University of Memphis, Memphis, Tennessee 38152.
2Center for Sport Medicine, The Pennsylvania State University, University Park, Pennsylvania 16802.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1998.
Publicado 22 de septiembre de 2004
Resumen
Palabras clave: fuerza muscular, testosterona, cortisol, hormona del crecimiento, péptido f
INTRODUCCION
El incremento en el volumen de entrenamiento y/o en la intensidad, que resulta en la reducción del rendimiento físico, ha sido definido como sobreentrenamiento (12, 14, 33). La fisiología del sobreentrenamiento ha sido estudiada para una variedad de actividades físicas (14, 33), siendo aquellas que enfatizan altos niveles de fuerza o potencia igual de susceptibles al sobreentrenamiento que otras actividades (14). En general, los mecanismos fisiológicos del sobreentrenamiento se han esclarecidos recientemente. Aunque queda mucho por estudiar acerca de la etiología del sobreentrenamiento, cuando se comparan actividades anaeróbicas, tales como los ejercicios de sobrecarga, con actividades aeróbicas, esto parece ser bastante diferente (12, 13). Muchos de los síntomas del sobreentrenamiento que han sido identificados para los ejercicios aeróbicos, tales como las alteraciones en los patrones del sueño, la frecuencia cardiaca de reposo, los estados de ánimo y la actividad simpática, no se han reportado para el sobreentrenamiento anaeróbico (12, 13). Además, la alteración de algunas de las variables agudas del entrenamiento para los ejercicios de sobrecarga (i.e., volumen de ejercicio, intervalos de recuperación y carga) producen una variedad de respuestas fisiológicas diferentes (30). De esta manera es probable que el sobreentrenamiento producido por programas de entrenamiento de la fuerza, claramente diferentes, pueda estar caracterizado por diferentes respuestas fisiológicas.
El incremento en los volúmenes de entrenamiento aeróbico (3, 34) y de entrenamiento de la fuerza (11, 18, 19) ha resultado en una reducción de la concentración de reposo de varias hormonas. Investigaciones recientes acerca del incremento en la intensidad del entrenamiento de la fuerza (i.e., porcentaje del máximo) (12, 13) indican que este tipo de estrés de entrenamiento podría ser fisiológicamente diferente del provocado por los incrementos en el volumen del entrenamiento de la fuerza. Pocos estudios acerca del entrenamiento de la fuerza han demostrado verdaderas disminuciones del rendimiento (34); así el sobreentrenamiento no está presente en muchas investigaciones. En general, pocos datos están disponibles acerca de las respuestas endocrinas al sobreentrenamiento provocado por el entrenamiento de la fuerza y las disminuciones concomitantes provocadas por el mismo.
Investigaciones previas acerca de la respuesta endocrina al incremento del estrés producido por el entrenamiento de la fuerza han monitoreado solamente las concentraciones hormonales de reposo (18). Se ha propuesto que las respuestas endocrinas inducidas por el ejercicio reflejan diferentes mecanismos de regulación hormonal (14), por lo cual recientes investigaciones han incluido las respuestas hormonales inducidas por el ejercicio (11, 17, 19). Los cambios en las concentraciones hormonales circulantes han sido propuestos como indicadores del sobreentrenamiento (1, 14), aunque dichos cambios hormonales no son siempre observados. Debido a que las fluctuaciones del rendimiento en la fuerza han sido asociadas con alteraciones en los perfiles endocrinos de reposo (16, 18) e inducidos por el ejercicio (27) en un proceso de entrenamiento de la fuerza, es posible que los cambios en las concentraciones hormonales puedan servir como marcadores de un inminente estado de sobreentrenamiento.
Recientemente, se ha desarrollado un protocolo de entrenamiento de la fuerza para producir disminuciones en la fuerza muscular a corto plazo con un bajo volumen (i.e., series x repeticiones), y una intensidad relativa alta [i.e., porcentaje de una repetición máxima (% 1 RM)] (12, 13). Aunque no fue pensado como un método de entrenamiento para alcanzar un rendimiento óptimo, este protocolo fue específicamente diseñado para provocar una respuesta de sobreentrenamiento, lo cual permite el estudio fisiológico del desarrollo de este fenómeno. Por ello, el propósito de esta investigación fue determinar si el sobreentrenamiento debido a una alta intensidad relativa (% de 1 RM) del entrenamiento de la fuerza provoca alteraciones en las concentraciones hormonales circulantes de reposo y en las inducidas por el ejercicio.
METODOS
Sujetos
Diecisiete hombres fueron aleatoriamente asignados a un grupo experimental que fue sobreentrenado mediante el uso altas intensidades relativas en el entrenamiento de sobrecarga (OT; n=11), o a un grupo control que utilizó bajas intensidades relativas de entrenamiento (Con; n=6). Todos los sujetos habían realizado entrenamiento de pesas para las piernas por un promedio de 4.5 años, y eran capaces de realizar el ejercicio de sentadilla con un peso equivalente a 1.5 veces el peso corporal, habían estado entrenando permanentemente antes de la participación en esta investigación, y no tenían historia de utilización de anabólicos esteroides durante el año previo al comienzo del estudio. Cada sujeto fue informado acerca de los riesgos de la participación y firmaron una declaración de consentimiento. Las características de estos mismos sujetos habían sido descritas previamente en estudios que examinaron la reducción del rendimiento y las respuestas de las catecolaminas al sobreentrenamiento provocado por entrenamiento de la fuerza de alta intensidad (12, 13). Las características de los sujetos de ambos grupos están presentadas en la Tabla 1.
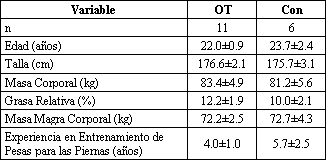
Tabla 1. Características de los sujetos de los grupos sobreentrenamiento y
control durante la evaluación 1. Los valores son presentados como medias±EE:
n, número de hombres. OT, grupo sobreentrenamiento, Con, grupo control. Los
valores no cambiaron significativamente durante el curso de la investigación,
p>0.05.
Protocolos para Entrenamiento de la Fuerza y para las Evaluaciones
El entrenamiento de las extremidades inferiores fue realizado en una máquina para sentadillas (Southern Xercise, Cleveland, TN). Las características cinemáticas de la sentadilla en esta máquina particular indican que este ejercicio no es idéntico a la sentadilla realizada con la barra, pero en cambio involucra principalmente la extensión de la cadera y de las rodillas (9). Este dispositivo fue elegido para este estudio, debido a que utiliza un patrón motor general fácil de aprender que involucra a grandes grupos musculares en una actividad multiarticular dentro de un rango de movimiento articular regulado. Se mejoró la seguridad de los sujetos debido a que una única persona podría ayudar fácilmente a los sujetos en los levantamientos máximos en el caso de una repetición fallida. Esta máquina también ha sido utilizada en estudios longitudinales previos acerca del entrenamiento de la fuerza (12, 13). En el presente estudio el entrenamiento para el tren inferior estuvo controlado, mientras que el programa de entrenamiento de la fuerza para el tren superior se mantuvo constante a lo largo de todo el período de entrenamiento. Todos los sujetos participaron en una fase de familiarización de 1 semana. Dos sesiones de la semana de familiarización fueron utilizadas para que los sujetos adquirieran la técnica apropiada de ejercicio y para valorar la fuerza en 1 RM. La confiabilidad de la evaluación de 1 RM, determinada a partir de las dos sesiones de familiarización fue de r=0.96. Las sesiones de ejercicio en la maquina para sentadillas fueron realizadas durante las semanas 2 y 3 del estudio. En cada día de entrenamiento el grupo OT realizó en la máquina de sentadillas, un protocolo de ejercicios con un bajo volumen y una alta intensidad relativa (% de 1 RM); se ha mostrado que esto provoca una respuesta de sobreentrenamiento caracterizada por reducciones en la fuerza muscular (12, 13). A la inversa, el grupo Con realizo un protocolo de entrenamiento con un bajo volumen y una baja intensidad relativa 1 día/semana, diseñado para mantener la fuerza muscular y no provocar un síndrome de sobreentrenamiento.
Tanto el ejercicio de sobrecarga, como el protocolo de evaluación de la fuerza utilizado en la presente investigación han sido descritos en detalle previamente (12, 13) y son presentados en la Figura 1. Brevemente, el objetivo de cada sesión de entrenamiento del grupo OT fue realizar ejercicios de sobrecarga a la máxima intensidad relativa (100% de 1 RM), a través de la realización de 10 series de 1 repetición al 100% de su RM evaluada en la maquina de sentadillas con 2 minutos de pausa entre cada intento de levantamiento. Cuando un peso no podía ser levantado, la sobrecarga para las repeticiones subsiguientes era reducida en 4.5 kg. El resultado neto fue que cada uno de los 10 levantamientos fue realizado tan cerca de la capacidad máxima del sujeto como fuera posible. Cada sesión de entrenamiento comenzaba con una entrada en calor controlada, y era seguida por una evaluación de 1 RM a partir de la cual se basaban las cargas subsiguientes para ese día. Este protocolo de entrenamiento no estuvo destinado a imitar los programas tradicionales de entrenamiento utilizados para optimizar el rendimiento en la fuerza, sino que fue una herramienta utilizada para provocar la respuesta de sobreentrenamiento con la concomitante reducción en el rendimiento. El grupo control realizo una sesión semanal de entrenamiento, lo que permitió mantener tanto la familiarización con el ejercicio de una manera no estresante como también la fuerza muscular a lo largo del estudio. Un segundo día a la semana estuvo destinado a la evaluación de la 1 RM en el grupo control. Todas las sesiones de entrenamiento estuvieron supervisadas por al menos uno de los miembros del equipo de investigación.
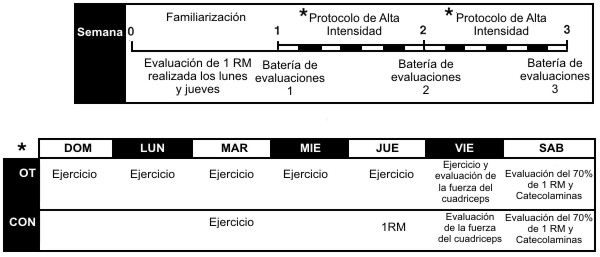
Figura 1. Línea de tiempo de las sesiones de ejercicio y de las sesiones de
evaluación para el grupo sobreentrenamiento (OT) y para el grupo control
(Con). Batería de evaluaciones 1, comienzo de la 2 semana del período de
sobreentrenamiento; batería de evaluaciones 2, mediados de la 2 semana del
período de sobreentrenamiento; Batería de evaluaciones 3, finales de la 2
semana del período de sobreentrenamiento. 1 RM, 1 repetición máxima. * hace
referencia a detalles en la parte inferior de la figura.
Las baterías de evaluaciones fueron administradas al comienzo (evaluación 1), a mediados (evaluación 2) y a finales de la segunda semana del período de sobreentrenamiento (evaluación 3). Durante la administración de cada batería de evaluaciones, se realizaron series de 10 repeticiones continuas hasta el agotamiento con el 70% de la 1 RM del sujeto en el momento de la evaluación, con 1 minuto de pausa entre cada serie. Este test sirvió como estímulo en el cual se monitorearon las concentraciones hormonales de reposo y de ejercicio; por ello, para minimizar las variaciones endocrinas diurnas, se mantuvo constante el momento del día en que cada sujeto fue evaluado.
Recolección de las Muestras de Sangre
Cuando se presentaron para la realización de la evaluación al 70% de 1 RM, los sujetos fueron colocados en posición de sentados en una posición similar a la posición final en la maquina de sentadillas. En la vena antecubital superficial se les insertó un catéter de teflón de 3.2 cm con un gauge de 20. La cánula se mantuvo regulada con un goteo continuo de solución salina (0.9% de cloruro de sodio) a una tasa de ~45 ml/hora. En línea con el goteo salino se colocó una llave de paso de cuatro vías para permitir la extracción de muestras sanguíneas con jeringas. Un ayuno de 6 hs precedió esta sesión de evaluación, aunque se les permitió a los sujetos beber agua ad libitum. Los sujetos permanecieron tranquilamente en la posición de sentados durante 30 minutos. Durante este tiempo, se extrajeron muestras de sangre luego de 15 (Pre 1) y 30 minutos (Pre 2). Esto fue inmediatamente seguido por el test realizado al 70% de 1 RM. Inmediatamente después de haber completado el test al 70%, se recolecto una muestra de sangre post ejercicio para la determinación de la concentración de péptido F. Los sujetos fueron entonces sentados durante 5 min, luego de los cuales se tomó una muestra final de sangre (5 min) para la determinación de las restantes concentraciones hormonales inducidas por el ejercicio.
Preparación de las Muestras de Sangre
Diez mililitros de sangre total de cada muestra fueron dejados coagular a temperatura ambiente y entonces fueron centrifugados a 1500g y 4ºC durante 15 minutos. El suero resultante fue dividido en alícuotas para los subsiguientes análisis de testosterona total, testosterona libre, cortisol y hormona del crecimiento. La concentración de péptido F fueron determinadas a partir de una muestras de 3 ml de sangre total que fueron inmediatamente colocadas en tubos de ensayo enfriados (4ºC) que contenían EDTA (3 mg) y aprotinina (1500 unidades inactivantes de calicreina. El plasma para el análisis del péptido F fue divido en alícuotas y luego fue centrifugado durante 15 min a 1500g a 4ºC. Un mililitro de sangre total fue mezclado con heparina y sodio, a partir del cual una pequeña porción fue utilizada para determinar el hematócrito por medio de la utilización de técnicas microcapilares estándar. La porción restante fue guardada para los subsiguientes análisis de la hemoglobina total. Todas las alícuotas no analizadas fueron inmediatamente guardadas a -90ºC para futuros análisis. Las muestras fueron descongeladas solamente una vez para el análisis y fueron decodificadas después que se hubieron completado todos los análisis (formato ciego).
Análisis Sanguíneos
Se utilizó el radioinmunoensayo de fase sólida con un único anticuerpo para determinar niveles de inmunoreactividad indicativos de concentraciones sanguíneas de testosterona, testosterona libre (Diagnostic Products, Los Angeles, CA) y cortisol (Diagnostic Systems, Webster, TX). La radioinmunoreactividad fue mediada con un contador y reductor de datos gama automatizado LBK Clinigamma 1272 (Pharmacia LKB Nuclear, Turku, Finlandia). Las varianzas para los radioinmunoensayos realizados en esta investigación fueron las siguientes: testosterona total (intra ensayo≤2.5%; inter ensayo=3.0%), testosterona libre (intra ensayo ≤2.5%, inter ensayo=9.6%), y cortisol (intra ensayo≤3.8%, inter ensayo=3.2%). Las concentraciones hormonales fueron determinadas en cada muestra tomada en el tiempo, para el cálculo de los índices testosterona/cortisol y testosterona libre/cortisol. El porcentaje de testosterona no unida fue determinado a partir los datos de la testosterona y de la testosterona libre para cada muestra tomada en el tiempo. Para la determinación de las concentraciones de hormona del crecimiento se utilizo el ensayo inmunoradiométrico (Nichols Institute Diagnostics, San Juan Capistrano, CA). Todas las muestras para la hormona del crecimiento fueron analizadas en duplicado con una varianza intra ensayo ≤3.7% y una varianza inter ensayo del 3.0%. El péptido F fue separado del plasma utilizando una columna separadora C18 (200 mg) (Peninsula Laboratories, Belmont, CA) y una serie de lavados (ácido trifluoroacético al 0.1% y acetonitrato al 60% en de ácido trifluoroacético al 0.1%). El eluyente resultante (3 ml) fue entonces evaporado a seco por un concentrador centrífugo. Las concentraciones de péptido F fueron entonces medidas con radioinmunoensayo de 125I, de fase líquida con doble anticuerpo, de 3 días (Peninsula Laboratories). Todas las muestras fueron analizadas en duplicado con una varianza intra ensayo ≤6.1% y una varianza inter ensayo del 14.0%. La recuperación del péptido F fue del 85.9%, y el coeficiente de regresión para las concentraciones estándar fue 0.991. Los valores de péptido F fueron entonces comparados con concentraciones de epinefrina previamente reportadas (13). Se calculó el índice molar entre el péptido F circulante y las concentraciones de epinefrina para cada muestra tomada en el tiempo. Los cambios en el índice molar a lo largo del tiempo sugerirían diferentes patrones de secreción para el péptido F y para la epinefrina. La hemoglobina total de toda la sangre fue determinada a través de calorimetría cuantitativa a 540 nm (Sigma Diagnostics, St. Louis, MO) utilizando un espectrofotómetro visible (Pharmacia LKB Novaspec II, Uppsala, Suecia). Los cambios en el volumen plasmático (%) inducidos por el ejercicio fueron estimados a partir de los cambios en el hematócrito y la hemoglobina utilizando los métodos de Dill y Costill (6).
Análisis Estadísticos
Todos los datos están presentados como medias±DE. Para comparar las características descriptivas de cada grupo se utilizó la prueba t para muestras independientes. El rendimiento físico fue analizado con un modelo mixto de ANOVA 2 x 3 (grupo x tets). Las variables sanguíneas fueron analizadas con un modelo mixto de ANOVA 2 x 3 x 3 (grupo x tiempo x test). Todos los análisis ANOVA fueron ajustados para muestras de diferentes tamaños. Se realizaron análisis post hoc con el procedimiento de la menor diferencia significativa de Fisher. La utilización de estadística paramétrica para todas las variables endocrinas dependientes fue respaldada por la falta de significacia estadística tanto para el test de distribución normal de Kolmogorov-Smirnov, como para el test de homogeneidad entre varianzas de Bartlett. El nivel de significación se estableció a una p<0.05 para todos los análisis.
RESULTADOS
Durante el curso de la investigación, las características descriptivas no cambiaron de los valores presentados en la Tabla 1 tanto para el grupo OT como para el grupo Con. Como se reportó previamente (12), la fuerza en 1 RM en el grupo OT se redujo significativamente en ~11% en comparación con el grupo Con (media del grupo OT=-12.2 kg; media del grupo Con =-1.1 kg). El grupo Con no exhibió cambios significativos en la fuerza en 1 RM, lo que fue indicativo de la eficacia de su protocolo de entrenamiento para mantener la fuerza específica de entrenamiento (1 RM).
Los resultados para la testosterona total, la testosterona libre, el porcentaje de testosterona no unida y el cortisol (Figura 2); el índice testosterona/cortisol y el índice testosterona libre/cortisol (Figura 3); para la hormona del crecimiento (Figura 4); y para el péptido F y su índice molar con la epinefrina (Figura 5), están ilustrados en sus respectivas figuras. No se evidenció una respuesta anticipadora (Pre 1 vs Pre 2) para ninguna de las variables en ningún punto en el tiempo. La testosterona, la testosterona libre y la hormona del crecimiento exhibieron un incremento significativo inducido por el ejercicio para la mayoría de las evaluaciones en ambos grupos. En el test 3, el grupo OT tuvo concentraciones de testosterona total post ejercicio ligeramente elevadas, mientras que el grupo Con en el test 3 exhibió concentraciones incrementadas tanto en reposo como post ejercicio. No se observaron cambios en el porcentaje de testosterona no unida a lo largo del tiempo para ninguno de los grupos o entre los grupos en algún punto en el tiempo. Tampoco se observaron cambios en las concentraciones de reposo de cortisol en ninguno de los grupos, y el ejercicio no produjo incrementos significativos, aunque en el test 3, el cortisol post ejercicio en el grupo OT disminuyó ligeramente.
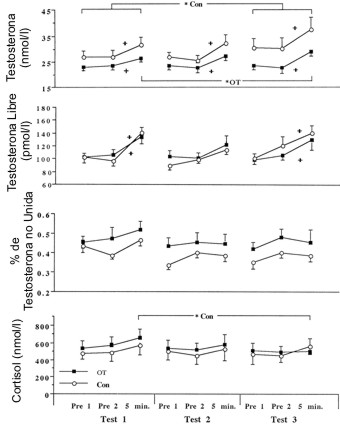
Figura 2. Respuesta en reposo e inducida por el ejercicio de la testosterona,
testosterona libre, porcentaje de testosterona no unida, y cortisol para ambos
grupos OT (n=11 hombres) y Con (n=6 hombres). Los valores son medias ± DE. Pre
1, 15 min pre ejercicio; Pre 2 inmediatamente antes del ejercicio; 5 min, 5
minutos post ejercicio. Los corchetes representan los puntos del tiempo
comparados. * Con y * OT, diferencia entre los test, P<0.05. + Diferente de
Pre 1 y Pre 2, P<0.05.
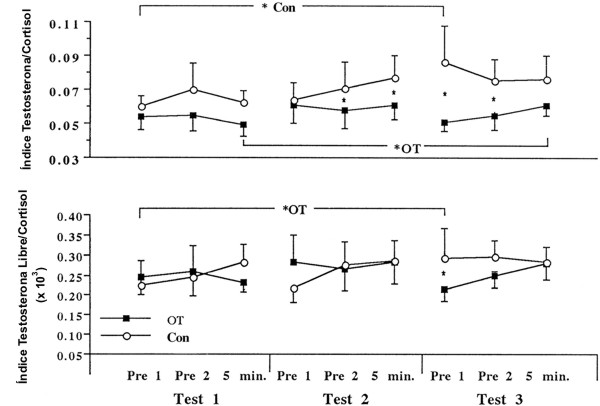
Figura 3. Respuesta en reposo e inducida por el ejercicio del índice
testosterona/cortisol y del índice testosterona libre/cortisol para ambos
grupos OT (n=11 hombres) y Con (n=6 hombres). Los valores son medias±DS. Los
corchetes representan los puntos del tiempo comparados. * Con y * OT,
representa diferencia entre los test, p<0.05. * representa diferencia entre
los grupos p<0.05.
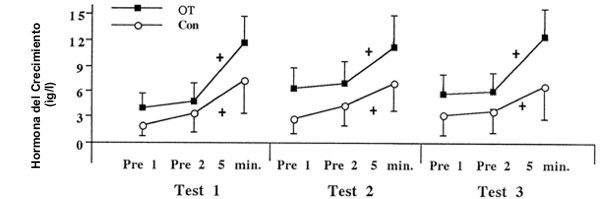
Figura 4. Respuesta de la hormona del crecimiento en reposo e inducida por el
ejercicio para ambos grupos OT (n=11 hombres) y Con (n=6 hombres). Los valores
son presentados como medias±DS. + Diferente con respecto a Pre 1 y Pre 2,
p<0.05.
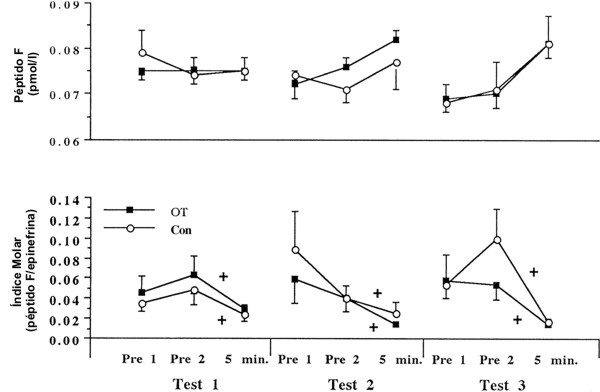
Figura 5. Respuesta del péptido F en reposo e inducida por el ejercicio y del
índice molar péptido F/epinefrina para ambos grupos OT (n=11 hombres) y Con
(n=6 hombres). Los valores son presentados como medias±DS. + Diferente con
respecto a Pre 1 y Pre 2; p<0.05.
La Figura 3 ilustra las respuestas de los índices testosterona total/cortisol y testosterona libre/cortisol. El índice testosterona total/cortisol post ejercicio se incremento ligeramente en el grupo OT en el test 3, aunque en este test, los valores pre ejercicio fueron menores que para los del grupo Con. Esta diferencia entre los grupos parece deberse a la elevada concentración de testosterona total en el grupo Con más que a una disminución en la testosterona total en el grupo OT. Se observaron pocas diferencias entre los grupos para el índice testosterona/cortisol. La hormona del crecimiento exhibió similares incrementos inducidos por el ejercicio para los tres test, pero no se observaron diferencias entre el grupo OT y el grupo Con. Las concentraciones de péptido F para el grupo OT y para el grupo Con fueron similares en todos los test, y no se reportaron cambios para ninguno de los grupos en ninguna de las muestras tomadas en el tiempo. El índice molar entre los valores de péptido F y los valores de epinefrina reportados previamente (13) no fue diferente entre los grupos OT y Con. En todos los test hubo una disminución significativa en el índice molar tanto en las muestras pre ejercicio (Pre 1 y Pre 2) como en las muestras tomadas inmediatamente post ejercicio. El cambio en el volumen plasmático medio estimado, inducido por el ejercicio, para todos los sujetos y todos los test fue de -15.9%. Esto indica que la no significación del incremento en el cortisol inducido por el ejercicio puede ser explicado por los cambios en el volumen plasmático. La testosterona total, la testosterona libre, y la hormona del crecimiento exhibieron incrementos mayores en la concentración inducida por el ejercicio de lo que puede atribuirse a cambios en el volumen plasmático por si solo. Aunque las concentraciones hormonales pueden se ajustadas a los cambios en el volumen plasmático (38), esto solo explica uno de los tantos mecanismos de regulación (30). Debido a que el interés del presente estudio es observar la concentración real de las hormonas circulantes y no la cinética hormonal, las concentraciones no fueron ajustadas para los cambios en el volumen plasmático, permitiendo por lo tanto la comparación con investigaciones previas acerca del sobreentrenamiento.
DISCUSION
Reducción en el Rendimiento
El protocolo de ejercicio para provocar el sobreentrenamiento utilizado en el presente estudio resulto en disminuciones significativas en la fuerza de las piernas en 1 RM (12), lo que fue indicativo de un estado de sobreentrenamiento (14, 33). Este fenómeno no parece haber sido transitorio, ya que los sujetos en el grupo OT reportaron una incapacidad para retomar las cargas normales de entrenamiento hasta 8 semanas después del estudio, por lo que requirieron un largo período de recuperación. Como se reportó previamente, la diminución en el rendimiento en la fuerza en el grupo OT no parece deberse al daño muscular, indicado por los niveles séricos de creatinquinasa total o por la percepción de dolor muscular (12). La disminución en el rendimiento de la fuerza fue resultado del entrenamiento de alta intensidad realizado solamente con el tren inferior, ya que el entrenamiento para el tren superior se mantuvo a niveles constantes para todos los sujetos. Aunque el comienzo del sobreentrenamiento provocado por el aumento del volumen ha sido estudiado en atletas de resistencia (34), las investigaciones previas no han monitoreado los perfiles endócrinos que acompañan el comienzo del sobreentrenamiento provocado por el entrenamiento de la fuerza de alta intensidad relativa (% 1 RM). El perfil hormonal observado en el presente estudio fue marcadamente diferente del observado para otros tipos de sobreentrenamiento y en general no fue alterado por el presente protocolo de sobreentrenamiento.
Testosterona Total
Aunque el incremento en los volúmenes del entrenamiento de pesas generalmente resulta en una reducción de los niveles de testosterona total en reposo e inducidos por el ejercicio (11, 15, 28), el incremento en la intensidad relativa de entrenamiento realizado por el grupo OT no solo no alteró la concentración pre ejercicio de testosterona total y en realidad provocó un ligero incremento en la respuesta aguda. Por otro lado, el grupo Con exhibió un incremento en la concentración de testosterona total pre ejercicio en el test 3, lo cual ha sido observado antes con la disminución del estrés de entrenamiento (15). Los incrementos en las concentraciones de testosterona total fueron mayores que los cambios en el volumen plasmático, lo que sugiere que una incrementada secreción de testosterona y/o una reducción en el clearance fue responsable de las respuestas inducidas por el ejercicio. La respuesta de la testosterona inducida por el ejercicio aparentemente no está bajo la regulación del sistema hipotalámico-pituitario a través de la hormona luteinizante (19). En el presente estudio, los incrementos agudos en la testosterona no estuvieron acompañados por incrementos en las concentraciones de hormona luteinizante (observaciones no publicadas); aunque, estos datos son preliminares ya que no explican las características pulsátiles. La elevada actividad simpática inducida por el ejercicio reportada previamente para el grupo OT (13), contribuyó más probablemente al aumento de la respuesta aguda de la testosterona (7, 23). El aumento de la respuesta en la testosterona total inducida por el ejercicio con el sobreentrenamiento en el test 3 pudo haber sido parte de un intento fisiológico no exitoso para preservar la capacidad de fuerza muscular. Aunque no es claro cual es el mecanismo exacto, el volumen relativamente bajo del entrenamiento de la fuerza pudo haber impedido la disminución de la testosterona total, en forma contraria a lo que se ha observado en los estudios donde se incrementó el volumen de entrenamiento de la fuerza (11, 15, 28).
Testosterona Libre
Los programas normales de entrenamiento de la fuerza producen un incremento en las concentraciones de testosterona libre (2, 18), mientras que el incremento en los volúmenes de entrenamiento resulta en una atenuación de la testosterona libre de reposo (16, 18, 19) y un aumento en la respuesta aguda de la misma (19). Se ha teorizado que la testosterona libre es más sensible que la testosterona total, al estrés producido por el sobreentrenamiento (16). De manera contraria a esta sugerencia, en ningún momento el protocolo realizado por el grupo OT en el presente estudio afectó las concentraciones de testosterona libre en ningún tiempo de muestreo. Los incrementos en la testosterona libre inducidos por el ejercicio fueron comúnmente evidentes y permanecieron constantes a lo largo del estudio.
Porcentaje de Testosterona no Unida
No se observaron cambios en el porcentaje de testosterona no unida, la cual es una fracción de la testosterona total clasificada como testosterona libre. Esto indica que los cambios en las proteínas fijadoras de testosterona, tales como la globulina fijadora de hormonas sexuales (SHBG) y la albúmina, simplemente reflejaron los cambios en la testosterona total circulante. Se ha sugerido que el incremento en la testosterona libre ocurre con el ejercicio hasta el agotamiento para compensar la reducción en la concentración de testosterona total (1). Debido a que la testosterona no unida es la fracción de la testosterona total que esta disponible para la actividad biológica en los receptores blanco, este podría ser un método efectivo para mantener la actividad biológica de la testosterona en presencia de una reducción en la concentración de la testosterona total (28). Esto, sin embargo, no fue evidente en el presente estudio. La falta de cambio en el porcentaje de testosterona no unida no fue inesperada, ya que la asociación constante de la SHGB no cambia con una serie aguda de ejercicios (7).
Cortisol
Los programas normales a largo plazo de entrenamiento de la fuerza no producen cambios o reducen la concentración de cortisol en reposo (2, 18) e incrementan la concentración inducida por el ejercicio (29). Al contrario de los estudios en donde se incrementó el volumen de entrenamiento de la fuerza (11, 16, 18), el protocolo de sobreentrenamiento utilizado en el presente estudio no produjo cambios en las concentraciones pre ejercicio de cortisol. De particular interés es la reducción inducida por el ejercicio en la concentración de cortisol en el grupo OT en el test 3, similar a lo que se observa con el incremento en los volúmenes de entrenamiento (11). La respuesta del cortisol inducida por el ejercicio fue en todo momento comparable o menor a los cambios en el volumen plasmático, indicando una falta de respuesta corticoadrenal al estímulo de ejercicio y sugiriendo que la duración del ejercicio en el presente estudio pudo haber sido muy corta (4). Es improbable que la regulación hipotalámico-pituitaria a través de la hormona adrenocorticotrofina sea responsable de la atenuación en la respuesta del cortisol inducida por el ejercicio, debido a que el tiempo de retraso en la activación del cortisol por la hormona adrenocorticotrofina es mucho mayor que el tiempo utilizado en este estudio para el muestreo del cortisol (8). Esto es además respaldado por la ausencia de algún cambio en los niveles de la adrenocorticotrofina a lo largo del curso del presente estudio (observaciones no publicadas), aunque estos datos son preliminares ya que no explican las características pulsátiles. Existe evidencia acerca del control neural de la actividad de la corteza adrenal (22), proporcionando de esta manera un mecanismo regulatorio veloz de la respuesta del cortisol inducida por el ejercicio. No se sabe, sin embargo, si la alteración en la actividad de la corteza adrenal, de la regulación por el sistema nervioso simpático o algún otro mecanismo es responsable de la disminución en la respuesta del cortisol inducida por el ejercicio observada en el grupo OT.
Índices Hormonales
Se ha sugerido que el índice testosterona total/cortisol puede ser un marcador del sobreentrenamiento provocado por el entrenamiento de sobrecarga (18). El índice testosterona total/cortisol inducido por el ejercicio comúnmente disminuye post ejercicio (10, 11). Si el volumen del entrenamiento de la fuerza es incrementado, el índice testosterona total/cortisol de reposo e inducido por el ejercicio va a disminuir (10, 16, 18), aunque este patrón es revertido con el entrenamiento a largo plazo y la exposición previa a elevados volúmenes de entrenamiento (10). En el presente estudio no se observaron disminuciones en el índice testosterona total/cortisol, y las diferencias entre el grupo OT y el grupo Con para los índices de reposo se debieron principalmente a un aumento en la testosterona total en el grupo Con solamente. El índice de reposo de la testosterona biológicamente disponible (i.e., testosterona libre) y cortisol (índice testosterona libre/cortisol) no cambia con el entrenamiento normal de la fuerza o con el desentrenamiento (2). En el presente estudio, se observaron pocos cambios en los valores del índice testosterona libre/cortisol en cualquiera de los grupos. Aunque el grupo OT experimento una reducción en el rendimiento, en ningún momento se observaron diminuciones en los valores del índice testosterona libre/cortisol ≥ 30% o por debajo de 0.30 x 10-3, al contrario de los marcadores de sobreentrenamiento sugeridos por Adlercreutz y cols. (1). En contraste con los reportes para actividades de resistencia, ni el índice testosterona/cortisol, ni el índice testosterona libre/cortisol parecen ser marcadores efectivos del sobreentrenamiento provocado por el entrenamiento de la fuerza de alta intensidad.
Hormona del Crecimiento
Ninguna investigación previa acerca del sobreentrenamiento provocado por el entrenamiento de la fuerza ha monitoreado las respuestas de la hormona del crecimiento. Los presentes datos sugieren que la hormona del crecimiento no es influenciada por el sobreentrenamiento provocado por el entrenamiento de la fuerza de alta intensidad. Aunque Barron y cols. (3) han reportado una disfunción hipotalámico-pituitaria con el sobreentrenamiento provocado por el entrenamiento de la resistencia, y Urhausen y cols. (39) han reportado recientemente una respuesta atenuada de la hormona del crecimiento con sobreentrenamiento provocado por entrenamiento de resistencia, los presentes datos no indican una alteración en las concentraciones de la hormona del crecimiento que acompañan al sobreentrenamiento provocado por el entrenamiento de la fuerza de alta intensidad.
Péptido F
El péptido F [preproencefalina (107-140)] es secretado por las células cromafinicas de la medula adrenal y a menudo es secretado en respuesta el mismo estímulo de ejercicio que causa la liberación de epinefrina (31). Investigaciones previas han indicado que, aunque el entrenamiento crónico fraccionado de alta intensidad y el entrenamiento de la resistencia en corredores universitarios resultó en aumentos de las concentraciones agudas de epinefrina, las concentraciones agudas de péptido F fueron atenuadas, sugiriendo que la epinefrina y el péptido F no son liberados de manera equimolar en corredores de elite altamente entrenados en resistencia (31). Esto contrastó con lo observado en individuos desentrenados quienes exhibieron concentraciones agudas equimolares del péptido F y de la epinefrina (31). Esta diferencia en las concentraciones agudas circulantes sugiere una posible adaptación al entrenamiento en los mecanismos de producción y/o secreción de las células cromafinicas adrenales. El péptido F puede desempeñar un rol en la regulación de la respuesta inmune a través de la regulación de las células natural killers, de la activación de neutrófilos, y de la mejora de la coagulación (21, 35, 40). Dicho efecto estimulante sobre el sistema inmune puede contrarrestar los efectos inmunosupresivos de las catecolaminas y de los glucocorticoides (21), los cuales están elevados en varios tipos de sobreentrenamiento (13, 14, 33). Esta inmunosupresión puede explicar parcialmente porque el sobreentrenamiento esta asociado con el incremento en la tasa de infección (14, 33).
Ningún estudio previo ha registrado la respuesta del péptido F al sobreentrenamiento provocado por el entrenamiento de la fuerza. El incremento agudo en la epinefrina reportado previamente debido a este protocolo de sobreentrenamiento (13) no fue observado para el péptido F. Estos datos sugieren que este protocolo de sobreentrenamiento no provoca secreciones equimolares para el péptido F y la epinefrina, como lo indica la reducción significativa en los índices molares inducidos por el ejercicio. La enorme respuesta de la epinefrina al sobreentrenamiento provocado por el entrenamiento de la fuerza de alta intensidad (13) pudo haber simplemente sobrepasado la capacidad de las células cromafinicas adrenales para secretar péptido F. Esto también sugiere que el requerimiento para el péptido F bajo estas condiciones estresantes es menor que para la epinefrina. Como se ha reportado previamente (32), la ausencia de una respuesta aguda del péptido F podría deberse a un inadecuado estímulo de ejercicio (e.g., duración demasiado corta, inadecuada producción de trabajo), o podría deberse al hecho de que todos los sujetos tenían experiencia en el entrenamiento de la fuerza, lo cual pudo haber atenuado la respuesta aguda del péptido F, tal como se ha reportado en corredores de resistencia altamente entrenados (31). No se sabe si la falta de respuesta del péptido F se debe a un enlentecimiento en la producción del péptido F a partir de su molécula precursora en comparación con la epinefrina (26, 37), o que hay conjuntos de células cromafinicas que puedan contener diferentes tasas de péptido F y de epinefrina o secretar preferencialmente un compuesto (31, 32). Debe señalarse que la vida media (t1/2) para el clearance del péptido F plasmático luego de haber sido secretado por las células cromafinicas es ≥15 min (24), lo cual le da al péptido F un tiempo considerable para que interactué con el sitio principal de acción, en la circulación (35, 40). El tiempo de recolección de muestras inmediatamente después del ejercicio fue suficientemente rápido para evitar problemas de diferencias en el t1/2 de clearance entre el péptido F y la epinefrina, y fue apropiado para determinar la concentración pico del péptido F post ejercicio (31, 32). Debe señalarse que no hay cambios conocidos en la tasa de clearance del péptido F debido a la intensidad o modalidad del ejercicio (31).
Marcadores del Sobreentrenamiento
Investigaciones previas han intentado identificar marcadores endócrinos para el síndrome de sobreentrenamiento inminente o concurrente, tanto para actividades aeróbicas (1, 3, 34) como anaeróbicas (18). Debido a la falta de asociación entre las alteraciones endócrinas y las alteraciones en el rendimiento, el tipo de ejercicio empleado en el presente estudio no permite el uso de las alteraciones hormonales para monitorear el sobreentrenamiento inminente. El protocolo de ejercicio diario utilizado por el grupo OT (i.e., una alta intensidad relativa, bajo volumen) puede no producir una activación aguda de gran amplitud en la función de la corteza adrenal o testicular, tal como se ha demostrado con un protocolo similar de entrenamiento de la fuerza (17). Esto podría explicar parcialmente la falta de adaptaciones endocrinas al protocolo utilizado por el grupo OT. Por lo tanto, para evitar el sobreentrenamiento producido por el entrenamiento de la fuerza de alta intensidad a corto plazo en sujetos con experiencia en el entrenamiento de pesas, puede ser necesario monitorear otros sistemas fisiológicos, tales como la actividad del sistema nervioso simpático (13), o las características neuromusculares (12). Investigaciones previas acerca del entrenamiento de la fuerza han demostrado correlaciones positivas significativas entre los cambios en la testosterona libre (2, 18), en el índice testosterona/cortisol (19) o el índice testosterona libre/cortisol (2) y los cambios en el rendimiento físico tal como la fuerza isométrica del cuadriceps (2, 19), la fuerza en 1 RM en el envión (18) o el índice de aptitud física (5). Ninguna de estas correlaciones fue evidente en el grupo OT (observaciones no publicadas).
Conclusión
Las concentraciones circulantes de testosterona, testosterona libre, cortisol, hormona del crecimiento y péptido F no fueron mayormente afectadas por el protocolo de entrenamiento de alta intensidad utilizado por el grupo OT. Aunque las reducciones en el rendimiento en la fuerza fueron visibles, estos cambios no estuvieron acompañados por alteraciones en el ambiente hormonal en el grupo OT. Aparentemente el sobreentrenamiento producido por el entrenamiento de la fuerza a corto plazo, de alta intensidad relativa no puede ser monitoreado exitosamente por medio de las concentraciones hormonales circulantes.
Agradecimientos
Los autores extienden su aprecio a N. Travis Triplett, L. Perry Koziris, Joseph L Marsit, Femke van Borselen y J. Michael Lynch por su ayuda en la recolección de los datos y a Steven J. Fleck, Robert S Staron y Anne B. Loucks por la ayuda en los análisis hormonales.
Dirección para el pedido de reimpresiones
A. C. Fry, 135 Roane Field House, University of Memphis, Memphis, TN 38152.
Referencias
1. Barron, J. L., T. D. Noakes, W. Levy, C. Smith, and R. P. Millar (1985). Hypothalamic dysfunction in overtrained athletes. J. Clin. Endocrinol. Metab. 60: 803-806
2. Brandenberger, G., and M. Follenius (1974). Influence of timing and intensity of muscular exercise on temporal patterns of plasma cortisol levels. J. Clin. Endocrinol. Metab. 40: 845-849
3. Dill, D. B., and D. L. Costill (1974). Calculation of percentage changes in volume of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 37: 247-248
4. Fahrner, C. L., and A. C. Hackney (1998). Effects of endurance exercise on free testosterone concentration and the binding affinity of sex hormone binding globulin (SHBG). Int. J. Sports Med. 19: 12-15
5. Farrell, P. A (1985). Adrenocorticotropic hormone and exercise. In: Exercise Endocrinology, edited by K. Fotherby, and S. B. Pal. New York: de Gruyter, p. 139-156
6. Fry, A. C., T. A. Aro, J. A. Bauer, and W. J. Kraemer (1991). A kinematic comparison of three barbell squat variations and a squat simulating machine (Abstract). J. Appl. Sport Sci. Res. 5: 162
7. Fry, A. C., W. J. Kraemer, M. H. Stone, B. J. Warren, S. J. Fleck, J. T. Kearney, and S. E. Gordon (1994). Endocrine responses to over-reaching before and after one year of weightlifting training. Can. J. Appl. Physiol. 19: 400-410
8. Fry, A. C., W. J. Kraemer, M. H. Stone, B. Warren, J. T. Kearney, S. J. Fleck, and C. A. Weseman (1992). The effect of amino acid supplementation on testosterone, cortisol, and growth hormone responses to one week of intensive weightlifting. Int. J. Sport Nutr. 3: 306-322
9. Fry, A. C., W. J. Kraemer, F. van Borselen, J. M. Lynch, J. L. Marsit, E. P. Roy, N. T. Triplett, and H. G. Knuttgen (1994). Performance decrements with high intensity resistance exercise overtraining. Med. Sci. Exerc. Sports 26: 1165-1173
10. ry, A. C., W. J. Kraemer, F. van Borselen, J. M. Lynch, N. T. Triplett, L. P. Koziris, and S. J. Fleck (1994). Catecholamine responses to short-term, high-intensity resistance exercise overtraining. J. Appl. Physiol. 77: 941-946
11. Fry, R. W., A. R. Morton, and D. Keast (1991). Overtraining in athletes, an update. Sports Med. 12: 32-65
12. Hiddinga, H. J., D. D. Isaak, and R. V. Lewis (1994). Enkephalin-containing peptides processed from the proenkephalin significantly enhances the antibody-forming cell response to antigens. J. Immun. 152: 3748-3759
13. Holzwarth, M. A., L. A. Cunningham, and N. Kleitman (1987). The role of adrenal nerves in the regulation of adrenocortical function. Ann. NY Acad. Sci. 512: 449-464
14. Jezova, D., and M. Vigas (1981). Testosterone response to exercise during blockade and stimulation of adrenergic receptors in man. Horm. Res. 15: 141-147
15. Katzenstein, G. E., D. Lund, P. Schultz, and R. V. Lewis (1987). Target tissue distribution of the proenkephalin peptides F, E and B. Biochem. Biophys. Res. Commun. 146: 184-1190
16. Kilpatrick, D. L., R. V. Lewis, S. Stein, and S. Undenfriend (1981). Release of enkephalins and enkephalin-containing polypeptides from perfused beef adrenal glands. Proc. Natl. Acad. Sci. USA 77: 7473-7475
17. Kopin, I. J (1977). Catecholamine metabolism (and the biochemical assessment of sympathetic activity). Clin. Endocrinol. Metab. 6: 525-549
18. Koziris, L. P., A. C. Fry, W. J. Kraemer, M. H. Stone, J. T. Kearney, S. J. Fleck, J. Thrush, S. E. Gordon, and N. T. Triplett (1992). Hormonal and competitive performance responses to an over-reaching stimulus in elite junior weightlifters (Abstract). J. Appl. Sports Sci. Res. 6: 186
19. Kraemer, W. J (1992). Hormonal mechanisms related to the expression of muscular strength and power. In: Strength and Power in Sport, edited by P. V. Komi. London: Blackwell Scientific, p. 64-76
20. Kraemer, W. J., A. C. Fry, B. J. Warren, M. H. Stone, S. J. Fleck, J. T. Kearney, B. P. Conroy, C. M. Maresh, C. A. Weseman, N. T. Triplett, and S. E. Gordon (1992). Acute hormonal responses in elite junior weightlifters. Int. J. Sports Med. 13: 103-109
21. Kraemer, W. J., L. Marchitelli, S. E. Gordon, E. A. Harman, J. E. Dziados, R. Mello, P. N. Frykman, D. McCurry, and S. J. Fleck (1990). Hormonal and growth factor responses to heavy resistance exercise protocols. J. Appl. Physiol. 69: 1442-1450
22. Kraemer, W. J., B. Noble, B. Culver, and R. V. Lewis (1985). Changes in plasma proenkephalin peptide F and catecholamine levels during graded exercise in men. Proc. Natl. Acad. Sci. USA 82: 6349-6351
23. Kraemer, W. J., J. F. Patton, H. G. Knuttgen, C. J. Hannan, T. Kettler, S. E. Gordon, J. E. Dziados, A. C. Fry, P. N. Frykman, and E. A. Harman (1991). Effects of high-intensity cycle exercise on sympathoadrenal-medullary response patterns. J. Appl. Physiol. 70: 8-14
24. Kuipers, H., and H. A. Keizer (1988). Overtraining in elite athletes, review and directions for the future. Sports Med. 6: 79-92
25. Lehmann, M., H. Dickhuth, G. Gendrisch, W. Lazar, M. Thum, R. Kaminski, J. F. Aramendi, E. Peterke, W. Wieland, and J. Keul (1991). Training-overtraining. A prospective, experimental study with experienced middle- and long-distance runners. Int. J. Sports Med. 12: 444-452
26. Lett, E., S. Gangloff, M. Zimmermann, D. Wachsmann, and J.-P. Klein (1994). Immunogenicity of polysaccharides conjugated to peptides containing T- and B-cell epitopes. Infect. Immun. 62: 785-792
27. Livett, A. R., D. M. Dean, L. G. Whelan, S. Undenfriend, and J. Rossier (1981). Co-release of enkephalin and catecholamines from cultured adrenal chromaffin cells. Nature 289: 317-319
28. Schwab, R., G. O. Johnson, T. J. Housh, J. E. Kinder, and J. P. Weir (1994). Acute effects of different intensities of weight lifting on serum testosterone. Med. Sci. Sports Exerc. 25: 1381-1385
29. Urhausen, A., H. H. W. Gabriel, and W. Kindermann (1998). Impaired pituitary hormonal response to exhaustive exercise in overtrained endurance athletes. Med. Sci. Sports Exerc. 30: 407-414
30. Vlahos, C. J., and W. F. Matter (1992). Signal transduction in neutrophil activation phosphatidylinositol 3-kinase is stimulated without tyrosine phosphorylation. FEBS Lett. 309: 242-248
31. Phillips, J. H (1987). Chromaffin granule biogenesis and the exocytosis/endocytosis cycle. In: Stimulus-Secretion Coupling in Chromaffin Cells, edited by K. Rosenheck, and P. I. Lelkes. Boca Raton, FL: CRC Press, vol. I, p. 32-49