
Respuestas Neuromusculares ante Cargas de Fuerza Explosiva y de Fuerza de Alta Intensidad
Paavo V Komi1, Keijo Häkkinen1, Robert U Newton2, V. Linnamo1, Allan Davie3, M. McGuigan3 y Travis Triplett-McBride4
1Neuromuscular Research Center and Department of Biology of Physical Activity, University of Jyväskylä, SF-40700 Jyväskylä, Finland.
2Human Perfomance Laboratory, Ball State University, Muncie, Indiana, U.S.A.
3School of Exercise Science and Sport Management, Southern Cross University, Lismore, New South Wales, Australia.
4Department of Exercise and Sport Science and The Department of Biology, University of Wisconsin – La Crosse
Artículo publicado en el journal PubliCE, Volumen 0 del año 2000.
Publicado 13 de octubre de 2003
Resumen
Palabras clave: fatiga neuromuscular, concéntrico, isométrico, espectro de potencia del EMG, composición del tipo de
INTRODUCCIÓN
El trabajo muscular intenso conduce a la fatiga, la cual puede depender del tipo de levantamiento (18, 21), la cantidad y la intensidad de la carga (17, 24), la composición de fibras rápidas/lentas del músculo ejercitado (40), y la especificidad de los antecedentes atléticos (16). La fatiga inducida por el ejercicio conduce a una disminución de la fuerza acompañada por disminuciones en la actividad electromiográfica medida de los músculos en condiciones voluntarias máximas (17, 18, 24), aunque los incrementos momentáneos en la actividad EMG máxima durante las muy rápidas, primeras y repetidas contracciones máximas son también posibles en algunas situaciones (4). La fatiga también conduce a un desplazamiento en el espectro de potencia del EMG hacía menores frecuencias (2, 19). Las frecuencias del espectro de potencia del EMG están relacionadas a la velocidad de conducción promedio de las fibras musculares activas (2) y la velocidad de conducción de las fibras musculares es más alta en las fibras de contracción rápida (1). Hay evidencia acerca de que un alto número relativo de fibras de contracción rápida resulta en valores de frecuencia más altos del espectro de potencia del EMG (11, 26) y que el desplazamiento durante la fatiga es generalmente mayor en los sujetos que poseen un mayor número relativo de fibras de contracción rápida (19, 42). El incremento de la concentración de lactato sanguíneo post-ejercicio parece estar relacionado al tipo y cantidad de carga (15, 17, 24), a la duración de los períodos de trabajo y descanso (31), así como al número relativo de fibras de contracción rápida (38, 43). Durante el ejercicio explosivo con pequeños períodos de trabajo y una velocidad de movimiento máxima la fatiga puede ser más de origen central, mientras que durante ejercicios de fuerza de alta intensidad la fatiga puede ser central y periférica (24).
Además de los estudios de fatiga, el espectro de potencia del EMG ha sido también usado para estudiar los cambios en la activación muscular a diferentes velocidades de movimiento. Ha sido sugerido que el desplazamiento del espectro de potencia del EMG hacia frecuencias más altas con velocidades de movimiento más altas ocurre debido a una diferencia en los patrones de activación con un mayor uso de unidades motoras rápidas (29, 37). Sin embargo, cuando se compara acciones dinámicas de diferentes intensidades, los cambios en las frecuencias de potencia promedio y en las frecuencias medianas varían entre sujetos (10). Además de la velocidad de movimiento, el tipo de acción puede tener un efecto sobre el patrón de activación. En la situación submáxima, durante las acciones excéntricas las unidades motoras rápidas podrían ser preferentemente reclutadas, mientras que en las acciones concéntricas las unidades motoras lentas pueden ser reclutadas primero y luego seguidas por las unidades motoras rápidas (30).
Para examinar si más acciones concéntricas intensas causarían algún cambio en los patrones de activación fueron elegidos dos ejercicios de piernas intensos, pero de diferente tipo. Además de observar diferentes velocidades de movimiento, también fue de interés estudiar los efectos de fatiga de los dos ejercicios. Las altas velocidades de movimiento, como las usadas en los ejercicios explosivos, podrían causar un desplazamiento del espectro de potencia del EMG hacia frecuencias más altas en comparación con las velocidades de movimiento bajas, si las unidades motoras rápidas están siendo preferentemente reclutadas. Este debería ser el caso durante la fatiga en los ejercicios explosivos, en los cuales es esperada una disminución en la frecuencia mediana debido a que las unidades motoras rápidas también se fatigan fácilmente. De otro modo, la fatiga periférica más substancial en los ejercicios de fuerza de alta intensidad pude conducir a una disminución en la velocidad de conducción de las fibras musculares, así causando una disminución en la frecuencia mediana.
El propósito de este estudio fue examinar (1) los efectos agudos de ejercicios concéntricos de prensa de piernas de fuerza explosiva y de fuerza de alta intensidad sobre el espectro de potencia del EMG, aEMG, producción de fuerza, y la concentración de lactato sanguíneo y (2) ver como estos cambios están relacionados a la distribución del tipo de fibras.
MATERIALES y MÉTODOS
Sujetos
8 hombres jóvenes adultos se ofrecieron como sujetos para el estudio. La edad, peso y talla medias (±DS) fueron 27±0.7 años, 74.4±3.2 kg, y 181.3±1.1 cm. Los sujetos eran físicamente activos y realizaban activamente varias actividades físicas, pero no tenían antecedentes de entrenamientos de fuerza regulares o deportes competitivos de ningún tipo. Fueron dadas advertencias acerca de posibles riesgos y e incomodidades a los sujetos y todos dieron su consentimiento por escrito para participar. El estudio fue conducido de acuerdo a la declaración de Helsinki y fue aprobado por el Comité Ético de la Southern Cross University.
Diseño del Experimento
El experimento fue de diseño transversal para evaluar dos ejercicios diferentes de press de piernas: 1) Ejercicio de fuerza explosiva (EE) y 2) Ejercicio de fuerza de alta intensidad (HRE). Los dos ejercicios fueron realizados separadamente de modo que hubo un período de recuperación de al menos dos semanas después de los ejercicios explosivos, los cuales fueron administrados primero. A los sujetos se les permitió continuar sus actividades físicas normales a través de todo el período del experimento, pero los mismos fueron instruidos para tener un día completo de descanso precediendo a las sesiones de evaluación.
Ejercicio de Fuerza Explosiva (EE)
El protocolo EE consistió de 5 x 10 repeticiones de extensiones concéntricas de piernas bilaterales en una máquina de prensa de piernas inclinada (Colosal, Sydney) con una ángulo de cadera de 110º. Los sujetos extendían sus piernas a partir de una posición de inicio de un ángulo de rodilla de 100º hasta la extensión completa, tan rápido como fuera posible. El sistema de frenado controlado por computadora detenía a la prensa y la bajaba (44), de modo que los sujetos no realizaran ningún trabajo excéntrico. Durante el EE el peso correspondió al 40±6% del máximo isométrico a un ángulo de rodilla de 100º. Fue elegido un ángulo de inicio de la rodilla grande de modo que los sujetos fueran capaces de realizar acciones explosivas de manera efectiva. El período de recuperación entre las series fue de un minuto.
Ejercicio de Fuerza de Alta Intensidad (HRE)
El HRE fue realizado usando el mismo protocolo y aparato que el EE y el número de series y repeticiones fue el mismo en ambos ejercicios. Sin embargo, en el HRE fueron adicionados pesos extras, de modo que las cargas para la primera serie correspondieran a cerca del 70% (67±7) del máximo isométrico a un ángulo de rodilla de 100º. Durante el HRE los pesos fueron, ya sea adicionados o quitados de modo que los sujetos fueran siempre capaces de justo finalizar las 10 repeticiones requeridas.
Mediciones
La fuerza isométrica máxima y la fuerza durante las acciones concéntricas fueron determinadas usando una plataforma de fuerza Kistler (Tipo 9287, Kistler, Suiza) la cual estaba adherida a la prensa de piernas. Fueron realizadas una extensión isométrica bilateral máxima de la rodilla seguida de 3 acciones concéntricas individuales con cargas submáximas de 40%, 55% y 70% del máximo isométrico a un ángulo de rodilla de 100º. Fue administrada una evaluación isométrica de fatiga a un nivel de fuerza del 10% de la contracción voluntaria máxima (MVC), manteniendo la fuerza en la intensidad objetivo requerida, lo cual era proyectado en la pantalla de la computadora. La fuerza objetivo fue incrementada cada 10 segundos, en incrementos de 10% hasta que el sujeto no era capaz de seguir manteniendo el nivel de fuerza requerido. Un período de trabajo de 10 segundos era siempre seguido de un período de descanso de 5 segundos. Todas las acciones isométricas fueron realizadas con un ángulo de rodilla de 120º.
La fuerza fue registrada en un sistema de computadora (Amlab software, Amlab Internacional, Australia) con una frecuencia de toma de muestras de 1 kHz. Fueron calculados el pico de fuerza máximo (N), fuerza promedio (N) y la máxima tasa de producción de fuerza (N/s) en acciones isométricas y dinámicas.
Un interruptor rotatorio (Omron Corporation, Japón) el cual estaba unido a la prensa y unido con una interfase a la computadora, medía la posición del aparato de prensa de piernas. Esto nos permitió calcular el tiempo de movimiento.
La actividad electromiográfica (EMG) fue registrada de los músculos vasto medial (VM) de ambas piernas con módulo de electrodos plata/cloruro de plata. Cada módulo de electrodo consistió de dos electrodos activos y un tercer electrodo de referencia, todos equidistantes a 2 cm. Los electrodos activos fueron alineados paralelos con las fibras a mitad de distancia entre el centro estimado de la zona de inervación y el tendón distal. La posición de los electrodos fue luego cuidadosamente marcada sobre la piel para asegurar la misma posición de los electrodos en ambos ejercicios. Antes de la aplicación de los electrodos, cada sitio fue afeitado y limpiado con alcohol para reducir la impedancia entre cada par de electrodos. Fueron incorporados preamplificadores (Quantec, Brisbane, Australia) dentro de los módulos de los electrodos, con la señal siendo amplificada con amplificadores (Quantec, Brisbane, Australia) a un ajuste de filtro de paso bajo de 1 kHz y a un paso alto de 3 kHz. Las señales EMG fueron registradas con un sistema de software Amlab con una frecuencia de muestreo de 1 kHz. Para calcular la EMG promedio (aEMG) la señal fue totalmente rectificada y la amplitud promedio de la señal fue calculada en el período de tiempo elegido. En ambas condiciones (isométricas y concéntricas) la comparación fue hecha solo con la acción usada en el ejercicio. Así la condición por si misma constituía su propio control y referencia y siempre para el mismo ángulo articular (isométrica) o rango de movimiento (concéntrica). Esto fue para eliminar los efectos que pueden causar a la señal los movimientos del electrodo en relación a la zona de inervación. Para el espectro de potencia de la EMG para analizar el mismo rango de movimiento, fueron usadas ventanas de 128 y 256 puntos en las acciones concéntricas explosivas y de fuerza de alta intensidad, respectivamente, y una ventana de 1024 puntos en las acciones isométricas para la transformación Fourier (FFT) para obtener frecuencias medianas (MF) y frecuencias de potencia medias (MPF) (Mega Electronics, Kuopio, Finlandia). En las acciones dinámicas la ventana fue colocada al inicio del movimiento, y en las acciones isométricas máximas la ventana fue colocada en la fase de meseta de la fuerza pico. Durante la evaluación de fatiga isométrica la ventana fue colocada en el punto medio de cada período de trabajo de 10 segundos.
Una cánula fue insertada dentro de una vena antecubital de cada sujeto antes del ejercicio. La sangre fue extraída antes de los ejercicios, y después de la segunda, cuarta y última serie en ambos ejercicios y analizada usando un analizador de lactato YSI 1500 Sport L-lactate Analyzer (Yellow Springs Instrument Co, Ohio, Estados Unidos).
Las biopsias musculares fueron obtenidas del músculo vasto lateral de seis sujetos por medio de un procedimiento estándar que involucro un método de succión (3, 9). Las fibras musculares fueron luego alineadas, montadas en pequeñas partes de corcho, congeladas en isopentano pre enfriado en nitrógeno líquido, y almacenadas a -80º para los análisis posteriores. Fueron realizados análisis histoquímicos estándar para determinar la distribución del tipo de fibras (6, 12). Las muestras fueron seccionadas transversalmente en serie (10 μm) y montadas en portaobjetos. Las muestras fueron analizadas para actividad de la mATPasa después de la preincubación ya sea a un pH de 4.34 (1.5-2 minutos, 25 ºC) o a un pH de 10.3 (5-7 minutos, 37 ºC). Además, otras muestras fueron también teñidas para NADH-tetrazolium reductasa (13). Las muestras fueron luego montadas en portaobjetos (Aquamount, BDH Laboratories, Poole, England) y fotografiadas (Olympus BH Imaging System, Olympus America Inc., Melvilla, Estados Unidos). La clasificación de fibras fue realizada manualmente contando aproximadamente 100 fibras (90-153) para cada muestra y luego remitiendo el pH 4.34, pH 10.3 y las muestras teñidas de NADH para determinar la proporción de fibras tipo I a tipo II.
Métodos Estadísticos
Fueron usados métodos estadísticos convencionales para los cálculos de medias, errores estándar (ES) y coeficientes de correlación. En las acciones concéntricas el promedio de la 2da, 3ra y 4ra repeticiones de la primera serie, y el promedio de las 4ta, 5ta y 6ta repeticiones de las 2da, 3ra y 4ta series y el promedio de la 7ma, 8va y 9na repeticiones de la quinta serie fueron usados para análisis posteriores. Los datos fueron analizados usando análisis de varianza múltiple (MANOVA). Cuando fue apropiado, fueron realizadas comparaciones de medias por medio de test t de Student apareados.
RESULTADOS
El tiempo promedio de una repetición concéntrica fue 347 ms en el EE y 670 en el HRE, y las fuerza correspondientes fueron 1121 N y 1556 N, respectivamente. La potencia promedio fue aproximadamente 39% más alta durante el EE que durante el HRE. Durante el HRE el tiempo promedio de una repetición concéntrica se incrementó en un 11.6±4.4% (P<0.05) desde la primera hasta la última serie, mientras que no hubo ningún cambio durante el EE (Tabla 1). No fueron observados cambios significativos en la fuerza pico, máxima tasa de producción de fuerza o fuerza promedio desde un ángulo de rodilla de 100º hasta 180º durante las acciones concéntricas de ambos ejercicios (Tabla 1).
Durante las acciones concéntricas del EE, a medida que el ejercicio progresaba, hubo un desplazamiento del espectro de potencia del EMG hacia frecuencias más altas. En comparación con la primera serie el valor MF más alto en el EE fue encontrado en la cuarta serie (P<0.05), después disminuyó ligeramente (Figura 1). Durante el HRE, la MF fue menor en comparación con el EE y disminuyo ligeramente (NS) hacia el final del ejercicio, de modo que la diferencia fue significativa (P<.005) comparada con la serie cinco del EE (Figura 1). No fueron encontrados cambios significativos en la aEMG entre ninguna serie tanto desde un ángulo de rodilla de 100º a 120º o de 100º a 180º (Figura 2).
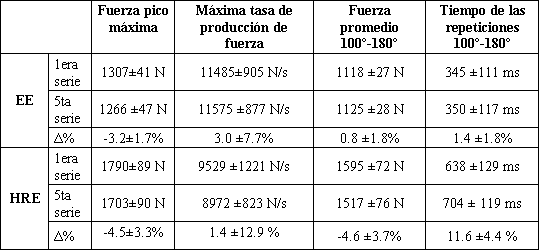
Tabla 1. Características de fuerza (±DS) durante las acciones
concéntricas del ejercicio de fuerza explosiva (EE) y fuerza de alta intensidad
(HRE) y sus cambios relativos (Δ%).
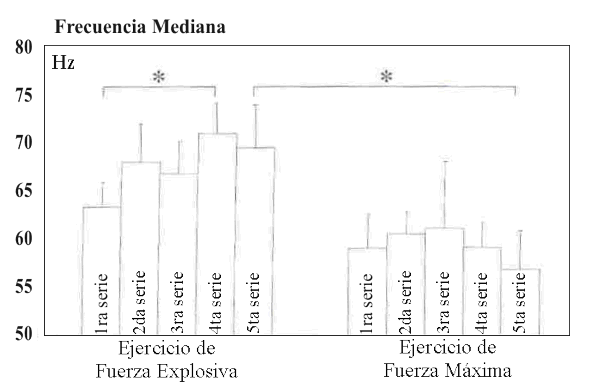
Figura 1. Frecuencia mediana (±DS) durante el ejercicio de fuerza
explosiva y fuerza de alta intensidad.
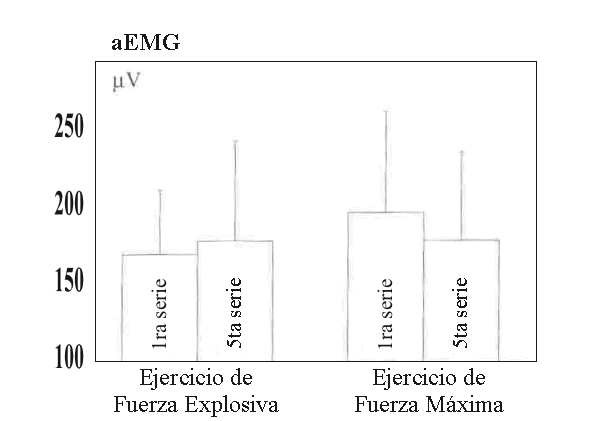
Figura 2. aEMG (±DS) durante la primera y quinta serie del ejercicio de
fuerza explosiva y fuerza de alta intensidad.
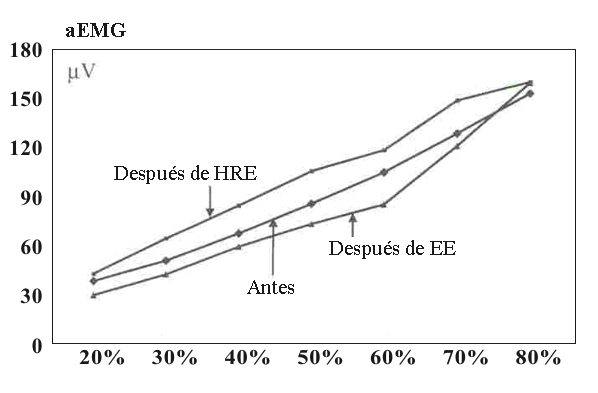
Figura 3. aEMG durante la evaluación isométrica de fatiga antes y después
del ejercicio de fuerza explosiva y fuerza de alta intensidad. La curva antes
representa el promedio de ambas curvas EE y HRE, las cuales no eran
significativamente diferentes una de la otra.
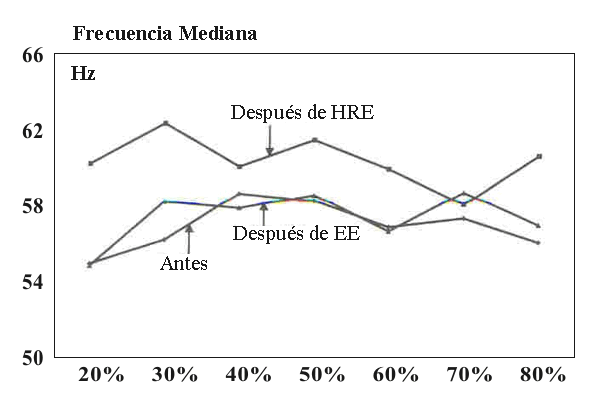
Figura 4. Frecuencia mediana durante la evaluación isométrica de fatiga
antes y después del ejercicio de fuerza explosiva y de fuerza de alta
intensidad. La curva antes representa el promedio de ambas curvas EE y HRE, las
cuales no eran significativamente diferentes una de la otra.
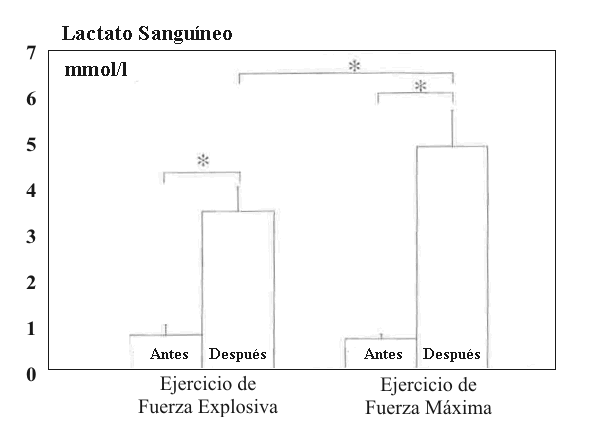
Figura 5. Lactato sanguíneo (±DS) antes y después del ejercicio de fuerza
explosiva y fuerza de alta intensidad.
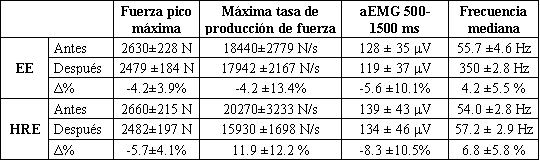
Tabla 2. Características de fuerza, aEMG (500-1500 ms) y MF en acciones
isométricas máximas (±DS) antes y después del ejercicio de fuerza explosiva (EE)
y el ejercicio de fuerza de alta intensidad (HRE) y sus cambios relativos (Δ%).
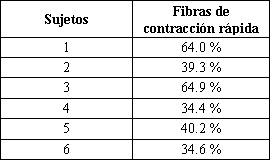
Tabla 3. Porcentaje de fibras de contracción rápida en cada sujeto.
No fueron encontradas diferencias significativas, en ya sea acciones concéntricas submáximas individuales o en la acción isométrica máxima para la fuerza pico, máxima tasa de producción de fuerza, aEMG, MF y MPF en el ejercicio o entre los ejercicios. La Tabla 2 resume los cambios en las acciones isométricas. En la evaluación de fatiga isométrica la EMG y MF fueron casi iguales antes y después del ejercicio en cada nivel de porcentaje de MVC durante el EE. Lo mismo fue cierto para el HRE, aunque fue observada una tendencia incremental (NS) (Figuras 3 y 4).
El lactato sanguíneo se incremento desde 1.00±0.22 hasta 3.09±0.55 mmol/l durante el EE (P<0.05) y desde 0.79±0.09 hasta 4.95±0.81 mmol/l durante el HRE (P<0.01). El incremento durante el HRE fue significativamente mayor (P<0.05) que durante el EE (Figura 5) y estuvo significativamente correlacionado con el cambio en la MPF en el HRE (r =-0.73; P<0.05).
El porcentaje de fibras rápidas varió desde 34.4% hasta 64.9% (Tabla 3). El número relativo de fibras de contracción rápida estuvo significativamente correlacionado con el cambio en el lactato sanguíneo en el HRE (r =0.87; P<0.05), mientras que el mismo no fue significativo en el EE (r =0.70; P<0.118). No fueron encontradas correlaciones significativas entre el número relativo de fibras de contracción rápida y la MF o MPF, antes y después de cualquiera de los ejercicios, aunque la MF fue algo menor después del HRE (r =-0.72; P<0.105).
DISCUSIÓN
Los presentes datos demuestran que los signos de fatiga durante ambas cargas fueron mucho menores de lo que pudiera ser esperado en base a otro estudio, entre ejercicios de extensiones de piernas de fuerza máxima y fuerza explosiva con el mismo número de series y repeticiones (24). Durante el EE hubo dificultosamente algún cambio o en algunas ocasiones un pequeño incremento en los parámetros de fuerza, y aún en el HRE las disminuciones no fueron grandes. Lo mismo también puede ser observado en el incremento en la concentración de lactato sanguíneo en el HRE, la cual fue extremadamente baja en comparación a estudios previos sobre protocolos de fuerza de alta intensidad (15, 22, 24, 39). Aunque el protocolo de series x repeticiones 5x10 RM fue previamente usado (24), los pequeños cambios en el HRE pueden ser explicados por (1) un pequeño rango de movimiento y consecuentemente una corta duración de tiempo de acción, (2) falta de trabajo excéntrico y (3) por la prensa de piernas de resistencia constante usada en este estudio.
Durante la fatiga ha sido observado un desplazamiento del espectro de potencia del EMG hacia frecuencias más bajas, en una situación isométrica (e.g., [2]) y en una situación dinámica (e.g., [19]). Los datos durante el HRE apoyan estos hallazgos, mientras que durante el EE en vez de una disminución, la MF y la MPF se incrementaron verdaderamente. Ha sido hallado que el componente de frecuencia del espectro de potencia correlaciona bien con la velocidad de conducción de las fibras musculares (2), la cual es más alta en las unidades motoras rápidas (1). Anteriormente has sido sugerido que una disminución en la velocidad de conducción esta relacionada a la acumulación de protones [H+] (23) y ha sido demostrado que la concentración de lactato está correlacionada con la frecuencia de potencia promedio (38, 43). En el presente estudio fue también encontrada una correlación negativa entre el cambio en la MPF y el lactato sanguíneo durante el HRE. Parece posible, sin embargo, que la acumulación de protones o de lactato sanguíneo no son principalmente responsables de los cambios espectrales del EMG de superficie. En su lugar, ha sido sugerido un deterioro del acoplamiento excitación-contracción como una causa del cambio (41). Durante la situación isométrica, el cambio en la velocidad de conducción para generar cambios iguales en el espectro de potencia del EMG fue mucho mayor en la ausencia de fatiga que durante la misma y fue sugerido que los cambios en la forma de la onda de las fibras musculares individuales no es el único contribuyente del desplazamiento de los componentes de frecuencia de la EMG de superficie (15).
Ha sido demostrado que el incremento de la temperatura muscular afecta el componente de frecuencia del espectro de potencia (32). Fue encontrado que el efecto del incremento de la temperatura sobre la potencia máxima es máximo a las más altas velocidades de pedaleo en un cicloergómetro isokinético (34), aunque ha sido demostrado que la MPF disminuye debido al enfriamiento, pero permanece sin cambios con el calentamiento (14). El tiempo total de trabajo con los mismos períodos de descanso fue mayor en el HRE, por lo que la temperatura muscular podría haberse incrementado aún más durante el HRE que durante el EE. Sin embargo, en el HRE no fue observado ningún incremento en la MF o MPF, pero más bien una tendencia a una disminución. Puede ser posible, que además del incremento en la temperatura, hubo también un efecto de aprendizaje. Si los sujetos fueran capaces de activar unidades motoras rápidas adicionales en el curso del ejercicio, la MPF y MF también se incrementarían. Ya que no fueron observados cambios en la aEMG a través de ambos ejercicios, un incremento en la temperatura muscular puede ser una explicación más probable de porque la disminución en la MF y MPF durante el HRE no fue más substancial y las mismas se incrementaron verdaderamente durante el EE. Ya que la MF fue algo mayor durante la cuarta serie, es probable que hubiera disminuido aún más si hubieran sido usadas series adicionales.
Una velocidad de movimiento más alta puede ser acompañada por cambios en la activación desde músculos lentos a rápidos (27, 35), y un desplazamiento del espectro de frecuencia del EMG hacia frecuencias más altas (29, 37). Algunos estudios anteriores han demostrado que un alto número relativo de fibras de contracción rápida puede estar relacionado a altos valores de frecuencia del espectro de potencia del EMG (11, 26). En el presente estudio esta relación no pudo ser demostrada, posiblemente debido al bajo número de sujetos o a las diferencias en la colocación de los electrodos entre los sujetos. Los electrodos fueron colocados a mitad de distancia entre el centro estimado de la zona de inervación y el tendón distal de todos los sujetos, pero la estimación no estuvo basada en la estimulación eléctrica. De este modo es posible que la distancia entre la zona de inervación y el electrodo no fuera la misma en todos los sujetos, lo cual pudo afectar el espectro del EMG (33). Durante las acciones concéntricas, aunque las fuerzas fueron más altas durante el HRE que durante el EE, la MF y la MPF fueron más altas en el EE. Esto pudo ser debido al mayor uso de unidades motoras rápidas en la situación explosiva, como fue especulado en algunos estudios anteriores de diferentes velocidades de movimiento (7, 29, 37). En las acciones dinámicas es posible que los electrodos pudieran cambiar su ubicación en relación a la zona de inervación, lo cual puede afectar el espectro de potencia del EMG (33). En el presente estudio el rango de movimiento fue similar en todas las velocidades, por lo tanto los posibles efectos deberían ser similares en ambos ejercicios. Otro problema puede ser el bajo número de puntos de datos usados para la ventana FFT, especialmente en el EE, lo que puede causar problemas con la estacionalidad de la señal. Los coeficientes de variación expresados como porcentajes (CV%) (25) entre la segunda y tercera repeticiones de la primera serie fueron 8.7% para MF y 6.8% para MPF en el EE, y 9.9% para MF y 7.0% para MPF en el HRE. El bajo CV% para MF y MPF indicaría que aún ventanas FFT más cortas pueden ser usadas en el análisis de los movimientos rápidos. Esto debería ser, sin embargo, cuidadosamente evaluado con un mayor número de sujetos en el futuro.
El propósito de las acciones concéntricas individuales con cargas submáximas fue también examinar el efecto de la velocidad de movimiento sobre la MF y MPF. Posiblemente las repeticiones individuales no fueron suficientes para activar completamente el sistema neuromuscular, ya que no fueron observados cambios en la MF y MPF. Cuando se compara la MF y MPF de acciones concéntricas individuales entre los dos ejercicios no fueron observadas diferencias significativas, sugiriendo que el espectro de potencia del EMG puede ser usado aún en mediciones separadas por varias semanas. La idea de la evaluación isométrica de fatiga fue examinar si un umbral de fatiga neuromuscular, como fue hallado en el ciclismo (28) podría ser también determinado en la situación isométrica, y como sería afectado el mismo por dos levantamiento diferentes. No pudo ser determinado ningún umbral claro en la evaluación isométrica de fatiga y no fueron observados cambios notables antes y después de los levantamientos. En el EE no hubo mucho cambio ya sea en la aEMG o en la MF y MPF, antes y después de los levantamientos. En el HRE, sin embargo, fue observada una tendencia incremental de la aEMG y los componentes de frecuencia del espectro de potencia a niveles submáximos, sugiriendo que después de la fatiga fue necesitada una mayor activación muscular para alcanzar el mismo nivel de fuerza relativa que antes de la fatiga (4). En estudios anteriores durante niveles de fuerza incrementados gradualmente, ha sido hallado un incremento en la MPF y MF del espectro de potencia (20, 33), mientras que hay también estudios sin ningún cambio con el nivel de fuerza (32, 42). En el presente estudio no hubo diferencias significativas en la MF y MPF entre los niveles de fuerza, pero pudo ser vista una tendencia incremental arriba del 40% de la MVC. De acuerdo con De Luca et al. (8) cuando todas las unidades motoras han sido reclutadas en algún nivel submáximo la velocidad de disparo continua para producir la fuerza máxima. Después, ya que la velocidad de conducción promedio no se incrementa, los siguientes cambios en las frecuencias espectrales pueden no ser tan pronunciados (36). Elegir un período de trabajo bastante largo (10 s) para la determinación del umbral neuromuscular puede haber causado los efectos de fatiga dentro de cada paso de fuerza, así contraponiéndose a los resultados concernientes al posible incremento de la MF y MPF con el incremento de la fuerza.
En conclusión, los presentes cambios de fatiga inducidos por un ejercicio de fuerza de alta intensidad apoyan los hallazgos de los estudios anteriores. Sin embargo, los cambios de fatiga, como fueron medidos por los cambios en la fuerza, lactato sanguíneo y aEMG no fueron obvios, posiblemente debido al pequeño rango de movimiento y el relativamente corto tiempo de acción, la falta de trabajo excéntrico y la prensa de piernas de resistencia constante usada en este estudio. El ejercicio explosivo, de otro modo, pareció haber facilitado la función del sistema neuromuscular en vez de producir fatiga. La potencia de frecuencia mediana y media se incrementaron durante el ejercicio explosivo y fueron más altas durante los movimientos rápidos en comparación con los movimientos más lentos. De este modo, parece que durante los movimientos rápidos puede ocurrir un incremento en la activación de las unidades motoras rápidas o una disminución de la activación de las unidades motoras lentas.
Dirección para correspondencia: Vesa Linnamo, Tel.:+358-14-2602054; fax: +358-14-2602071. Correo electrónico: linnamo@pallo.jyu.fi
Referencias
1. Andearssen S, Aredt-Nielsen L (1987). Muscle fibre conduction velocity in motor units of the human anterior tibial muscle: a new size principle parameter. J Physiol; 391: 561-71
2. Arendt-Nielsen L, Mills K (1985). The relationship between mean power frequency of the EMG spectrum and muscle fiber conduction velocity. Electroencep Clin Neurophysiol; 60: 130-4
3. Bigland-Ritchie B, Furbusg F, Woods J (1986). Fatigue of intermittent submaximal contractions central and peripheral factors. J Appl Physiol, 61 (2): 421-9
4. Bigland-Ritchie B, Donovan E, Roussos C (1981). Conduction velocity and EMG power spectrum changes in fatigue of sustained maximal efforts. J Appl Physiol Respirat Environ Exerc Physiol, 51 (5): 1300-5
5. Brooke MH, Kaiser KK (1970). Three myosin ATPase systems: the nature of their pH lability and sulfhydryl dependence. J Histochem Cytochem; 18: 670-2
6. Critterio G, Agostini E (1984). Selective activation of quadriceps muscle fibers according to bicycling rate. J Appl Physiol, 57: 371-9
7. De Luca C, LeFever R, McCue M, Xenasis A (1982). Behavior of human motor units in different muscles during linearly varying contraction. J Physiol, 329: 113-28
8. Evans WJ, Phinney SD, Young VR (1982). Suction applied to a muscle biopsy maximizes simple size. Med Sci Sports Exerc; 14 (1): 101-2
9. Gamet D, Duchene J, Garapon-Bar C, Goubel F (1990). Electromyogram power spectrum during dynamic contractions at different intensities of exercise. Eur J Appl Physiol, 61: 331-7
10. Gerdle B, Wretling M-L, Henriksson-Larsen K (1988). Do the fibre-type proportion and the angular velocity influence the mean power frequency of the electromyogram?. Acta Physiol Scand; 134: 341-6
11. Green HJ, Reichmann H, Pette D (1982). A comparison of two ATPase based schemes for histochemical muscle fibre typing in various mammals. Histochemistry; 76: 21-31
12. Halkjaer-Kristensen J, Ingemann-Hansen T (1979). Microphotometric analysis of NADH-tetrazolium reductase and α-glycerophosphate dehydrogenase in human quadriceps muscle. Histochemical J; 11: 127-36
13. Holowijn M, Heus R (1992). Effects of temperature on electromyogram and muscle function. Eur J Appl Physiol; 65: 541-5
14. Komi PV, Rusko H (1974). Quantitative evaluation of mechanical and electrical changes during fatigue loading of eccentric and concentric work. Scand J Rehab Med Suppl, 3: 121-6
15. Komi PV, Tesch P (1979). EMG frequency spectrum, muscle structure and fatigue during dynamic contractions in man. Eur J Appl Phusiol; 42: 41-50
16. Komi PV, Vittasalo J (1976). Signal characteristics of EMG at different levels of muscle tension. Acta Physiol Scand, 96: 267-76
17. Komi PV, Vittasalo J (1987). Changes in motor unit activity and metabolism in human skeletal muscle during and after repeated eccentric and concentric contractions. Acta Physiol Scand, 100: 246-54
18. Kraemer W, Noble B, Clark M, Culver B (1987). Physiologic responses to heavy-resistance exercise with very short periods. Int J Sports Med, 8: 247-52
19. Madsen O (1996). Torque, total work, power, torque acceleration energy and acceleration time assessed on a dynamometer: reliability of knee and elbow extensor and flexor strength measurements. Eur J Appl Physiol, 74: 206-10
20. Moritani T, Gaffney F, Carmichael T, Hargis J (1985). Interrelationships among muscle fiber types, electromyogram, and blood pressure during fatiguing isometric contraction. In: Winter D, Norman R, Well R, Hayes K, Patla A, editors. Biomechanics IX-A. Champaign, IL: Human Kinetics Publishers, 287-92
21. Moritani T, Oddson L, Leonard C, Thorstensson A (1995). Changes in activation and H-reflex amplitudes within the ankle extensor synergy during different modes of locomotion. In: ISB XVth Congress of the International Society of Biomechanics, Jyväskylä. Book of Abstracts, 638-9
22. Moritani T, Takaishi T, Matsumoto T (1993). Determination of maximal power output at neuromuscular fatigue thershold. J Appl Physiol, 74 (4): 1729-34
23. Muro M, Nagata N, Moritani T (1983). Analysis of myoelectric signals during dynamic and isometric contractions. In :Biomechaniscs, VIII-A. Champaign, IL: Human Kinetics Publishers, 432-9
24. Nardone A, Romano C, Schieppati M (1989). Selective recruitment of high-thershold human motor units during voluntary isotonic lengthening of active muscles. J Physiol, 409: 451-71
25. Pattersson R, Pearson J, Fisher S (1985). Work-rest periods: their effects on normal physiologic response to isometric and dynamic work. Arch Phys Med Rehabil, 66: 348-52
26. Petrofsky J, Lind A (1980). The influence of temperature on the amplitude and frequency components of the EMG during brief and sustained isometric contractions. Eur J Appl Physiol, 44: 189-200
27. Roy S, DeLuca C, Schneider J (1986). Effects of electrode location on myoelectric conduction velocity and median frequency estimates. J Appl Physiol, 61: 1510-7
28. Sargeant A (1987). Effect of muscle temperature on leg extension force and short-term power output in humans. Eur J Appl Physiol, 56: 693-8
29. Smith J, Betts B, Edgerton V, Zernicke R (1980). Rapid ankle extension during paw shakes: Selective recruitment of fast ankle extensors. J Neurophysiol, 43: 612-20
30. Strojnik V, Jereb B, Colja I (1997). Median frequency change during 60 s maximal hopping and cycling. In: Second Annual Congress of the European College of Sport Sciences, Copenhagen, Denmark. Book of Abstracts I, 370-1
31. Tesch P (1980). Muscle fatigue in man with special referente to lactate accumulation. Acta Physiol Scand, 480 (Suppl): 1-40
32. Thorstensson A, Karlsson J (1976). Fatiguability and fibre composition of human skeletal muscle. Acta Physiol Scand, 98: 318-22
33. Vestergaard-Poulsen P, Thomsen C, Sinkjaer T, Henriksen O (1995). Simultaneous 31P-NMR spectroscopy and EMG in exercising and recovering human skeletal muscle: a correlation study. J Appl Physiol, 79 (5): 1469-78
34. Viitasalo J, Komi PV (1978). Isometric endurance, EMG power spectrum, and fiber composition in human quadriceps muscle. In: Asmussen E, Jorgensen K, editors. Biomechanics VIA. Baltimore: University Park Press, 244-50
35. Viitasalo J, Komi PV (1981). Effects of fatigue on isometric force- and relaxation-time characteristics in human muscle. Acta Physiol Scand, 111: 87-95
36. Wilson G, Newton R, Murphy A, Humphries B (1993). The optimal load for the development of dynamic athletic performance. Med Sci Sports Exerc, 25 (11): 1279-86