
Seis Sesiones de Entrenamiento Intervalado de Esprint Incrementan el Potencial Oxidativo del Músculo y la Capacidad de Resistencia en Ciclismo en Humanos
Martin J Gibala1, Kirsten A Burgomaster1, Scout C Hughes1, George J Heigenhauser2 y Suzanne N Bradwel1
1Exercise Metabolism Research Group, Department of Kinesiology, McMaster University, Hamilton, Ontario, Canadá.
2Department of Medicine, McMaster University, Hamilton, Ontario, Canadá.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2005.
Publicado 3 de octubre de 2008
Resumen
Palabras clave: citrato sintetasa, glucógeno muscular
INTRODUCCION
Realizar sesiones repetidas de ejercicio del tipo de “esprints” de alta intensidad a través de varias semanas o meses induce cambios profundos en el músculo esquelético. Han sido descritas un amplio intervalo de adaptaciones musculares metabólicas y morfológicas (25, 34); sin embargo, la magnitud y dirección de cambio en muchas variables depende de la naturaleza del protocolo de entrenamiento, i.e., la frecuencia, intensidad y duración de los esfuerzos de esprint, así como la recuperación entre sesiones. Dada la contribución significativa del metabolismo energético aeróbico durante los esprints repetidos (3, 26, 29, 40), no es sorprendente que haya sido reportado (19, 25) un incremento en el potencial oxidativo del músculo, tal como es indicado por los cambios en las actividades máximas de “marcadores” enzimáticos tales como la citrato sintetasa, luego de 6-8 semanas de entrenamiento de esprint. Recientemente, dos estudios reportaron grandes incrementos en la actividad máxima de la citrato sintetasa, así como en el consumo de oxígeno pico (VO2 pico), luego de solo 2 semanas de entrenamiento de esprint diario (30, 33). Estos datos sugieren que las mejoras en el metabolismo energético aeróbico pueden ser rápidamente estimuladas por medio de sesiones cortas de ejercicio muy intenso; sin embargo, se desconoce el efecto de menos sesiones de esprint. Además, aparte de los cambios en el VO2 pico, no estamos al tanto de datos que sugieran que el entrenamiento de esprint conduce a un incremento de la capacidad de realizar ejercicio que sea principalmente de naturaleza “aeróbica”, e.g., una prueba de resistencia hasta la fatiga a una carga submáxima fija.
De este modo, el propósito principal del presente trabajo es estudiar el efecto de seis sesiones de entrenamiento intervalado de esprint sobre el potencial oxidativo del músculo, el VO2 pico y el tiempo de resistencia hasta la fatiga durante el ciclismo a una intensidad equivalente a ~ 80% del VO2 pico. En base a un trabajo piloto en nuestro laboratorio que mostró mejoras modestas en el rendimiento luego de 6 días consecutivos de entrenamiento de esprint, decidimos emplear una intervención de 2 semanas, de modo que sean permitidos 1-2 días de descanso entre las sesiones de entrenamiento, en un esfuerzo de promover la recuperación y facilitar las adaptaciones de rendimiento. La importancia de los días de descanso entre las sesiones de entrenamiento fue enfatizada en un estudio reciente (30), que mostró que la potencia pico y media producida durante un test de Wingate no cambió después de 14 días consecutivos de entrenamiento de esprint; sin embargo, cuando los sujetos realizaron el mismo número de sesiones de entrenamiento a través de 6 semanas (i.e., con 1-2 días de descanso entre las sesiones de entrenamiento), la producción de potencia se incrementaba significativamente. Aunque numerosos mecanismos podrían estar potencialmente implicados, la importancia de los días de descanso entre las sesiones de entrenamiento puede estar relacionado en parte al hecho de que el entrenamiento intenso conduce a la inactivación de las bombas musculares de cationes (23, 36), y ha sido especulado que pueden ser requeridos varios días para la normalización de la función de la bomba de Ca2+ del retículo sarcoplásmico (41).
Así, el modo y la intensidad de los esfuerzos de esprint en el presente estudio fue similar al de dos estudios recientes que incorporaron intervenciones de entrenamiento de 2 semanas (30, 33), sin embargo, el volumen total fue reducido en aproximadamente dos tercios y en total implicó solo ~ 15 min de ejercicio a través de 2 semanas. Nosotros planteamos la hipótesis acerca de que nuestro protocolo de entrenamiento de esprint de corta duración incrementaría el potencial oxidativo del músculo y la capacidad de resistencia en el ciclismo. También medimos la concentración de glucógeno muscular en reposo, debido a que solo unos pocos estudios de entrenamiento de esprint han realizado esto, lo que ha producido resultados conflictivos (14, 27, 30, 33, 28).
Nuestro diseño experimental incluyó a un grupo control, que completó los test de rendimiento con ~ 2 semanas de separación sin ninguna intervención de entrenamiento, y todos los sujetos realizaron varias pruebas de familiarización antes de realizar las evaluaciones de la línea de base o condición inicial.
METODOS
Sujetos
Dieciséis individuos sanos se ofrecieron como voluntarios para participar en el experimento (Tabla 1). Ocho sujetos (2 mujeres) fueron asignados a un grupo de entrenamiento y realizaron tests de ejercicio antes y después de una intervención de entrenamiento de esprint de 2 semanas. Otros ocho hombres sirvieron como grupo control y realizaron el test de rendimiento ~ 2 semanas después sin ninguna intervención de entrenamiento. También obtuvimos muestras de biopsias con aguja a partir del grupo de entrenamiento para estudiar las potenciales adaptaciones inducidas por el entrenamiento en los músculos esqueléticos en reposo. No obtuvimos biopsias del grupo control, por razones éticas, debido a que otros estudios no han demostrado cambios en las concentraciones de metabolitos musculares en reposo o en las actividades máximas de enzimas mitocondriales cuando los sujetos control son evaluados con varias semanas de separación sin ninguna intervención de esprint (1, 28). Todos los sujetos eran recreacionalmente activos y pertenecían a la población estudiantil de la Universidad de McMaster, quienes participaban en alguna forma de ejercicio dos o tres veces por semana (e.g., trote, ciclismo, aerobics), pero ninguno estaba implicado en alguna forma de programa de entrenamiento estructurado. Luego de un chequeo médico de rutina, los sujetos fueron informados acerca de los procedimientos que iban a ser empleados en el estudio y de los riesgos asociados, y todos proporcionaron un consentimiento informado por escrito. El protocolo experimental fue aprobado por la Universidad de Mc Master y el Comité de Ética para la Investigación en Ciencias de la Salud de Hamilton.

Tabla 1. Características de los sujetos. Los datos son presentados como valores medios±DS para 8 sujetos. VO2 pico, consumo de oxígeno pico.
Procedimientos Pre-experimentales
Antes de formar parte de la prueba experimental (i.e., antes de las mediciones de línea de base o en condición inicial), todos los sujetos realizaron pruebas de familiarización para orientarse con todos los procedimientos de evaluación y dispositivos de entrenamiento. Específicamente, todos los sujetos realizaron: 1) un test de VO2 pico; 2) un “entrenamiento de práctica” para establecer la carga de trabajo que producía ~ 80% del VO2 pico; y 3) un test de capacidad de resistencia que consistió de ciclismo hasta la fatiga volitiva a ~ 80% del VO2 pico en por lo menos dos ocasiones separadas.
Detalles de los Tests de Rendimiento
Test de VO2 pico
Los sujetos realizaron un test incremental hasta el agotamiento en una bicicleta ergométrica frenada electrónicamente (Excalibur Sport V2.0, Lode, Groningen, Holanda) para determinar el VO2 pico usando un sistema de recolección de gases on-line (sistema modular de captación de oxígeno Moxus, tecnologías AEI, Pittsburg, PA). Los tres estadios iniciales del test consistieron de intervalos de 2 min a 50, 100 y 150 W, respectivamente, y la carga de trabajo fue incrementada en 25 W cada minuto hasta el agotamiento volitivo. El valor usado para el VO2 pico correspondió al valor más alto alcanzado a través del período de recolección de 30 s.
Test de Capacidad de Rendimiento en Ciclismo
Los sujetos pedalearon hasta el agotamiento volitivo en una bicicleta ergométrica frenada electrónicamente (Lode) a una carga de trabajo diseñada para provocar ~ 80% del VO2 pico. Todas las pruebas de rendimiento fueron conducidas en ausencia de retroalimentación temporal, verbal, o fisiológica. El test terminaba cuando la cadencia de pedaleo cayera debajo de 40 rpm (de acuerdo a las especificaciones del fabricante, la producción de potencia indicada por el equipo podría no haber sido válida por debajo de esta cadencia), también fue registrada la duración del ejercicio. Fueron recolectadas muestras de aire espirado para la determinación de la ventilación minuto, el consumo de oxígeno, producción de dióxido de carbono, e índice de intercambio respiratorio, los datos fueron promediados a través del período de ejercicio de 6 a 10 min.
Reproducibilidad de los Test de Rendimiento
Diez individuos que no fueron sujetos del presente estudio realizaron un test de VO2 pico y el test de capacidad de resistencia en ciclismo en días separados y por lo menos con una semana de separación, de modo que fue calculada la reproducibilidad de error del método, tal como fue descrito por Sale (35). El coeficiente de variación para el test de VO2 pico y el test de capacidad de resistencia en ciclismo fue de 3,7 y 12,0%, respectivamente.
Protocolo Experimental
El protocolo experimental consistió de 1) evaluaciones de línea de base o condición inicial (i.e., después de los procedimientos de evaluación descritos arriba); 2) una intervención de entrenamiento de esprint durante 2 semanas o período similar sin este tipo de entrenamiento (grupo control); y 3) post-evaluaciones, tal como se describe más abajo.
Evaluaciones de Línea de Base
Las mediciones de línea de base para todos los sujetos consistieron de un test de VO2 y un test de capacidad de resistencia en ciclismo. Cada test de línea de base fue conducido en un día separado con 24 h entre los tests. A los sujetos en el grupo de entrenamiento también se les realizó un procedimiento de biopsia muscular 3 días después del test de capacidad de resistencia de ciclismo en la condición inicial y varios días antes del inicio de la intervención de entrenamiento. Para los procedimientos de biopsia, el área sobre la porción lateral de un de los muslos fue anestesiada (lidocaína 2%, AstraZeneca Canada, Notario, Canadá) y fue realizada una pequeña incisión a través de la piel y de la fascia subyacente para permitir que sea obtenida una muestra de tejido (50-100 mg) del músculo vasto lateral (1). Los detalles respecto del protocolo experimental están resumidos en la Figura 1.
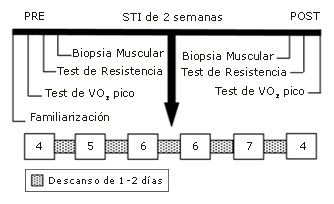
Figura 1. Descripción del protocolo experimental. VO2 pico, consumo de oxígeno pico; PRE, pre-ejercicio; POST, post-ejercicio; SIT, entrenamiento intervalado de esprint. Los números en las cajas indican el número de tests de Wingate completados durante cada una de las 6 sesiones de entrenamiento a través del período de 2 semanas.
Entrenamiento
El entrenamiento fue iniciado 3-5 días después de los procedimientos de biopsia de la condición inicial y consistieron de seis sesiones de entrenamiento intervalado de esprint distribuidos en 14 días. Cada sesión de entrenamiento consistió de esfuerzos máximos repetidos de 30 s en una bicicleta ergométrica frenada electrónicamente (Lode) contra una resistencia equivalente a 0,075 kg/kg de masa corporal (i.e., un test de Wingate). Los sujetos fueron instruidos para comenzar pedaleando tan rápido como fuera posible contra la resistencia inercial del ergómetro, ~ 2 s antes de que la carga apropiada fuera aplicada por una computadora unida mediante una interfase con un ergómetro y que tenía instalado el software apropiado (software Wingate versión 1.11, Lode). Los sujetos fueron alentados verbalmente para continuar pedaleando tan rápido como fuera posible a través del test de 30 s. La potencia pico, media y el índice de fatiga fueron determinados subsiguientemente usando un sistema de adquisición de datos on-line. Durante el período de recuperación de 4-min entre los tests, los sujetos permanecieron sobre sus bicicletas y descansaron o se les permitió pedalear a una cadencia baja (<50 rpm) contra una resistencia baja (<30 W) para reducir la acumulación venosa en las extremidades inferiores y minimizar las sensaciones de mareo o nauseas. El protocolo de entrenamiento consistió de un ejercicio realizado tres días por semana en días alternados (i.e., Lunes, Miércoles, Viernes) durante 2 semanas. El número de tests de Wingate realizados cada día durante el entrenamiento se incrementó desde 4 a 7 a través de las primeras cinco sesiones de entrenamiento, y en la sesión final, los sujetos completaron cuatro intervalos, tal como se resume en la Figura 1.
Post-evaluaciones
Fue obtenida una segunda muestra de biopsia muscular 3 días después de la sesión de entrenamiento final, para estudiar los cambios inducidos por el entrenamiento en el músculo en reposo, y fue iniciada una segunda batería de tests de rendimiento 2 días después de los procedimientos de biopsia (Figura 1). El grupo control realizó una segunda serie de tests ~ 2 semanas después de los tests de línea de base. La naturaleza de las mediciones de rendimiento de ejercicio post-evaluación fue idéntica en todos los aspectos a los tests de línea de base.
Control Dietario
En un intento de minimizar cualquier variabilidad potencial inducida por la dieta en el metabolismo del ejercicio y el perfil metabólico de reposo del músculo esquelético, se instruyó a los sujetos para que consumieran el mismo tipo y cantidades de alimentos durante las fases de condición inicial y post-evaluación. Los sujetos en el grupo de entrenamiento fueron particularmente alentados para mantener sus dietas de un modo tan similar como sea posible durante las 24 h antes de los procedimientos de biopsia pre- y post-entrenamiento. A los sujetos se les pidió que registraran toda la comida ingerida durante estos períodos, y la adherencia fue valorada realizando análisis dietarios sobre los registros individuales de alimentos mantenidos por los sujetos.
Los diarios de alimentación pre- y post-entrenamiento fueron analizados para ingesta energética total y proporción de energía derivada de los carbohidratos, grasas y proteínas (Nutritionist Five, First Data Bank, San Bruno, CA). Estos análisis confirmaron que no hubo ninguna diferencia entre las pruebas en la cantidad total de energía consumida o en las proporciones de macronutrientes.
Análisis Musculares
Una vez removida de la pierna, cada muestra de biopsia muscular fue inmediatamente congelada introduciendo la aguja de biopsia dentro de nitrógeno líquido. Las muestras fueron subsiguientemente divididas en dos partes mientras permanecían congeladas, y una pieza fue mantenida en nitrógeno líquido para la determinación de la actividad de las enzimas musculares. La parte restante de cada muestra fue deshidratada por congelación, espolvoreada y disecada para que estuviera libre de sangre y tejido conectivo, y fue almacenada a -85 °C antes de los análisis de metabolitos.
Citrato Sintetasa
Las muestras musculares húmedas fueron inicialmente homogeneizadas usando los métodos descritos por Henriksson y Reitman (17) hasta una dilución de 50 veces. La actividad máxima de la citrato sintetasa fue determinada en un espectrofotómetro (Ultrospec 3000 pro UV/Vis) usando un método descrito por Carter et al. (16). El coeficiente de variación intraensayo para el ensayo de citrato sintetasa, basado en 10 repeticiones de la muestra muscular, fue 4,9 %. El contenido de proteínas del homogenato fue determinado por el método de Bradford (5), usando un kit de ensayo comercial (Quick Stara, Bio-Rad Laboratorios, Hercules, CA), y los datos enzimáticos fueron expresados como moles por kilogramo de proteína por hora.
Metabolitos
Una alícuota de músculo deshidratado por congelación fue extraída en hielo usando ácido perclórico 0,5 M (que contenía 1 mM de EDTA), fue neutralizada con KHCO3 2,2 M, y el sobrenadante resultante fue usado para la determinación de todos los metabolitos, excepto glugógeno. El ATP, la fosfocreatina y la creatina fueron medidos usando ensayos enzimáticos adaptados para fluorometría (Hitachi F-2500, Hitachi Instruments, Tokio, Japón) (15, 31). Para el análisis de glucógeno, una alícuota de ~ 2 mg de músculo deshidratado por congelación fue incubada en HCl 2,0 N y calentada durante 2 h a 100 °C para hidrolizar el glucógeno hasta sus unidades glucosídicas. La solución fue subsiguientemente neutralizada con un volumen igual de NaOH 2,0 N y analizada para glucosa usando un ensayo enzimático adaptado para fluorometría (31).
El coeficiente de variación intraensayo para todos los ensayos de metabolitos musculares, basados en 10 repeticiones de la misma muestra, estuvieron entre un 2 y 3%. Todas las mediciones de metabolitos musculares fueron corregidas con la concentración total pico de creatina para un dado sujeto.
Análisis Estadísticos
Todos los datos de rendimiento de los ejercicios fueron analizados usando ANOVA para mediciones repetidas de dos factores. Para el test de Wingate, test de capacidad de resistencia y el test de VO2 pico, los factores fueron la prueba (pre-entrenamiento, post-entrenamiento) y la condición (entrenamiento, control). Para la comparación de la producción de potencia durante la primera vs. la última sesión de entrenamiento de esprint (solo el grupo de entrenamiento), los factores fueron la prueba (pre-entrenamiento, post-entrenamiento) y la serie de esprint (1-4). Todos los datos musculares fueron analizados usando test t de a pares (a dos colas). El nivel de significancia de los análisis fue establecido a un nivel p<0,05, y las interacciones significativas y los efectos principales fueron subsiguientemente analizados usando un test post hoc de diferencias significativas de Tukey. Todos los datos son presentados como valores medios±DS.
RESULTADOS
Capacidad de Resistencia en Ciclismo
Después del entrenamiento, las mejoras individuales en la capacidad de resistencia en ciclismo estuvieron entre un 81 y 169% en comparación con la línea de base, con la excepción de un sujeto (incremento de 16%), quien al final del estudio, reveló que había tenido una pequeña lesión en el hombro (no relacionada al experimento) el día antes de su prueba post-entrenamiento. Aún con la inclusión de los datos de este sujeto (Figura 2), el incremento medio en el tiempo hasta la fatiga en el test de resistencia de ciclismo fue de 100% para el grupo de entrenamiento (n=8) en comparación con la línea de base (51±11 vs. 26±5 min, p<0,05) y esto el incremento fue mayor (p<0,05) que el del grupo control, el cual no presentó cambios en el rendimiento (Figura 2). El consumo de oxígeno durante el ejercicio no fue diferente entre la primera y la segunda prueba en ninguno de los grupos; sin embargo, la ventilación minuto (post-entrenamiento: 91±7 vs. preentrenamiento: 104±9 L.min-1) y el índice de intercambio respiratorio (post-entrenamiento: 1,18 vs. pre-entrenamiento: 1,24) fueron menores (p<0,05) después del entrenamiento en el grupo de entrenamiento de esprint (p<0,05). El VO2 pico no cambió en ninguno de los grupos a través del transcurso del estudio.
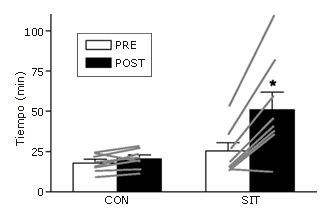
Figura 2. Tiempo hasta la fatiga en el test de resistencia de ciclismo antes y después de un protocolo de entrenamiento de esprint de 2 semanas (grupo de entrenamiento; SIT) o período equivalente sin entrenamiento (control; Con). Los datos son presentados como valores medios±DS. Los datos individuales también son graficados para todos los sujetos de cada grupo. * p<0,05.
Capacidad Anaeróbica de Trabajo
La producción de potencia pico durante cada uno de los cuatro tests de Wingate consecutivos realizados durante la última sesión de entrenamiento (sexta) fue mayor (p<0,05) en comparación con la primera sesión (Figura 3). Sin embargo, el índice de fatiga fue también más elevado (p<0,05) post-entrenamiento, y así no hubo diferencias en la producción de potencia media para cada uno de los cuatro tests de Wingate durante la primera en comparación con la última sesión de entrenamiento.
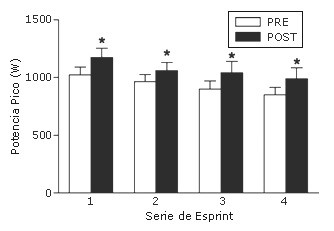
Figura 3. Potencia anaeróbica pico producida durante 4 tests de Wingate consecutivos realizados durante la primera y la última sesión de entrenamiento de esprint. Los datos son presentados como valores medios±DS.
Actividad de la Citrato Sintetasa y Concentraciones de Metabolitos Musculares en Reposo
La actividad máxima de la citrato sintetasa se incrementó (p<0,05) en un 38% después del entrenamiento (Figura 4). La concentración muscular de glucógeno en reposo se incrementó (p<0,05) en un 26% después del entrenamiento (Figura 5); sin embargo, no hubo cambios inducidos por el entrenamiento en las concentraciones musculares en reposo de ATP, fosfocreatina o creatina (Tabla 2).
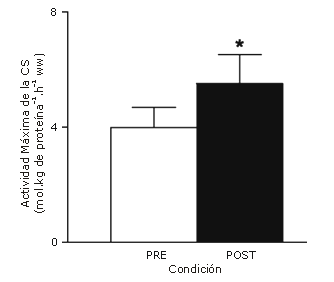
Figura 4. Actividad máxima de la citrato sintetasa (CS) medida en las muestras de biopsia muscular en reposo, obtenidas antes y después de un protocolo de entrenamiento de esprint de 2 semanas. Los datos son presentados como valores medios±DS para 8 sujetos, ww, peso húmedo. * p<0,05.
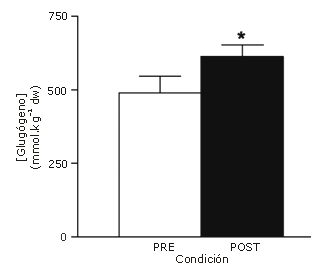
Figura 5. Concentración de glucógeno muscular medida en las muestras de biopsia en reposo obtenidas antes y después del protocolo de entrenamiento de esprint 2 semanas. Los datos son presentados como valores medios±DS para 8 sujetos, dw, peso seco. * p<0,05.

Tabla 2. Metabolitos musculares antes y después del entrenamiento. Los datos son presentados como valores medios±DS para 8 sujetos y son expresados en mmol.kg-1 de peso seco.
DISCUSION
El principal hallazgo original del presente estudio fue que seis sesiones de entrenamiento intervalado de esprint realizadas durante 14 días incrementan el potencial oxidativo y duplican el tiempo de resistencia hasta la fatiga durante el ciclismo a ~ 85% del VO2 pico en sujetos recreacionalmente activos.
La validez de esta última observación está reforzada por el hecho de que todos los sujetos realizaron pruebas de familiarización extensivas antes de las evaluaciones y el grupo control no presentó cambios en el rendimiento de resistencia cuando se lo evaluó 2 semanas después sin ninguna intervención de entrenamiento de esprint. También detectamos incrementos en el contenido de glucógeno muscular en reposo luego del entrenamiento de esprint. De este modo, los presentes datos demuestran que sesiones repetidas de esfuerzos de ciclismo máximo de 30 s, totalizando ~ 15 min de ejercicio total a través de 2 semanas, incrementaron dramáticamente la capacidad de resistencia en ciclismo y alteraron favorablemente el perfil metabólico de reposo del músculo esquelético humano. Aunque previamente han sido reportados incrementos en la actividad de la citrato sintetasa y del contenido de glucógeno después de varias semanas de entrenamiento intervalado de esprint (19, 25, 30, 33), los datos son equívocos (8, 10, 14, 24, 27) y nosotros demostramos aquí que el volumen total de entrenamiento necesario para estimular estas adaptaciones metabólicas es substancialmente menor que el que ha sido sugerido previamente.
Potencial Oxidativo del Músculo y Contenido de Glucógeno después del Entrenamiento Intervalado de Esprint de Corta Duración
Nosotros medimos la actividad máxima de la citrato sintetasa en biopsias de músculo esquelético en reposo antes y después del entrenamiento, debido a que esta constituye el marcador más comúnmente usado del potencial oxidativo del músculo, y otras investigaciones han justificado su selección de esta enzima, debido a que existe en una proporción constante con otras enzimas mitocondriales (e.g., Ref. 13). Existen datos equívocos respecto del efecto del entrenamiento de esprint sobre la actividad máxima de esta enzima; sin embargo, los estudios que han fallado en observar un incremento en la citrato sintetasa generalmente usaron esprints muy cortos, con una duración < 10 s (8, 24) o esprints que no constituyeron esfuerzos máximos (10). De manera contraria, todos los estudios que han reportado incrementos en la actividad de la citrato sintetasa incorporaron sesiones de esprint con esfuerzo máximo que duraron entre 15-30 s (19, 25, 30, 33).
Otra consideración relevante es el hecho de que el ejercicio agudo per se puede elevar la actividad de la citrato sintetasa, confundiendo potencialmente la interpretación de los efectos inducidos por el entrenamiento, y así el transcurso de tiempo del muestreo de músculo, relativo a la última sesión de ejercicio es crítico cuando se mide la actividad de esta enzima (22, 39). En el presente estudio, dejamos 72 h de recuperación antes de realizar cualquier procedimiento de muestreo con biopsia (i.e., después de la evaluación de línea de base y después de la sesión de entrenamiento final) para minimizar los efectos potenciales de confusión del ejercicio agudo anterior sobre la actividad de la citrato sintetasa (22).
Nuestros datos muestran claramente que la actividad máxima de la citrato sintetasa aumentó luego de solo seis sesiones de entrenamiento intervalado de esprint.
De manera notable, la magnitud del incremento fue similar a la reportada en otros estudios que incorporaron un número substancialmente mayor de sesiones de entrenamiento de esprint (19, 25, 30, 33).
Además, el incremento en la actividad de la citrato sintetasa en el presente estudio es comparable al reportado por algunos autores después de 6-7 días de entrenamiento de resistencia tradicional (i.e., 2 h/día a ~ 65% del VO2 pico) (7, 38), mientras que otros autores no han reportado cambio en el potencial oxidativo muscular después de entrenamiento de resistencia corto (e.g., Ref. 12). Los presentes datos no explican el mecanismo para la regulación en ascenso (upregulation) de la actividad de la citrato sintetasa, y están garantizados trabajos adicionales a este respecto. Finalmente, aunque hay datos limitados y equívocos respecto del efecto del entrenamiento intervalado de esprint sobre las reservas de glucógeno muscular en reposo (34), nuestros resultados son consistentes con dos estudios recientes que reportaron un incremento del contenido de glucógeno muscular después de 14 sesiones de entrenamiento intervalado de esprint (30, 33). Un aspecto particularmente nuevo de nuestros datos es que la magnitud del incremento en el glucógeno muscular fue comparable a lo que ha sido reportado después de cinco a siete sesiones de entrenamiento de resistencia tradicional (incremento promedio: ~ 20 %, intervalo: 13-35%; e.g., Refs. 7, 12, 32).
Efecto del Entrenamiento Intervalado de Esprint Corto sobre el Rendimiento de Resistencia
Varios estudios han reportado incrementos en el VO2 pico después de 14-24 sesiones de entrenamiento intervalado de esprint realizado durante 2-8 semanas (8, 10, 25, 26). Sin embargo, aparte de estas observaciones, no estamos al tanto de ningún dato que sugiera que el entrenamiento de esprint conduce a un incremento de la capacidad de realizar ejercicios que sean de naturaleza principalmente aeróbica. Así, en el presente estudio, decidimos emplear un test para valorar la capacidad de resistencia en el ciclismo a ~ 80% del VO2 pico, una tarea en la cual la vasta mayoría de la energía es aportada por el metabolismo oxidativo. Nuestros datos muestran que la capacidad de resistencia anaeróbica se incrementó dramáticamente después de solo seis sesiones de entrenamiento intervalado de esprint, a pesar del hecho de que el VO2 pico permaneció sin cambios.
Efectivamente, el tiempo de ejercicio hasta el agotamiento aumentó más del doble en seis de los ocho sujetos, quienes realizaron la intervención de entrenamiento (ver datos individuales en la Figura 2) y la mejora media en el rendimiento fue de 100%.
Parece improbable que este hallazgo sea un resultado falso, dado que el incremento medio fue substancialmente mayor que la variabilidad encontrada día a día para este test en nuestro laboratorio (coeficiente de variación = 12%) y a que el grupo control no mostró cambios en la capacidad de resistencia en ciclismo cuando fue evaluado ~ 2 semanas después de no realizar la intervención de entrenamiento de esprint. Para nuestro conocimiento, este es el primer estudio en demostrar que el entrenamiento intervalado de esprint mejora dramáticamente la capacidad de resistencia durante un test con carga fija, en el cual la mayor parte de la energía celular es derivada del metabolismo aeróbico.
Solo podemos especular sobre los potenciales mecanismos responsables del incremento dramático en la capacidad de rendimiento de resistencia, pero es plausible que un incremento en el potencial mitocondrial inducido por el entrenamiento, tal como fue medido por la actividad de la citrato sintetasa, haya mejorado la sensibilidad de control respiratorio durante el ejercicio, tal como ha sido clásicamente propuesto (18). Sin embargo, los mecanismos precisos que regulan el rendimiento de resistencia son multifactoriales y extremadamente complicados (9), y los datos de otros estudios sugieren que el entrenamiento de esprint puede estimular una serie de adaptaciones que podrían facilitar el rendimiento aparte de los cambios en el potencial mitocondrial.
Por ejemplo, investigaciones recientes han demostrado que el entrenamiento intervalado de esprint de 5-8 semanas incrementa el flujo sanguíneo y la conductancia muscular al músculo esquelético (21), la capacidad de transporte de lactato y la liberación de H+ desde el músculo activo (20), la regulación iónica (14) y la función del retículo sarcoplásmico (28). Aunque el transcurso de tiempo para que se produzcan estas adaptaciones es desconocido, otros estudios han reportado adaptaciones similares luego de solo 5-7 días de entrenamiento aeróbico, incluyendo cambios en la cinética de flujo sanguíneo (37), expulsión de lactato desde el músculo en ejercicio (4), y actividad de la bomba de cationes (11). Esperamos que las presentes observaciones estimulen a la realización de investigaciones adicionales para clarificar la naturaleza precisa, transcursos de tiempo, y significancia de las adaptaciones fisiológicas inducidas por el entrenamiento intervalado de esprint.
En conclusión, los resultados del presente estudio demuestran que seis sesiones de entrenamiento intervalado de esprint durante 2 semanas (~ 15 min totales de ejercicio muy intenso) incrementan la actividad máxima de la citrato sintetasa y duplican la capacidad de resistencia durante un ejercicio de ciclismo a ~ 80% del VO2 pico en sujetos recreacionalmente activos. La validez de esta última observación está reforzada por el hecho de que todos los sujetos realizaron pruebas de familiarización extensivas antes de las evaluaciones y a que un grupo control no mostró cambios en la capacidad de resistencia de ciclismo cuando fue evaluado 2 semanas después sin haber realizado la intervención de entrenamiento de esprint. Para nuestro conocimiento, este es el primer estudio que demostró que el entrenamiento de esprint incrementa dramáticamente la capacidad de resistencia durante un test con carga de trabajo fija, en el cual la mayor parte de la energía celular es derivada del metabolismo aeróbico. Estos datos demuestran que sesiones repetidas y breves de ejercicio de ejercicio muy intenso pueden estimular rápidamente mejoras en el potencial oxidativo muscular que son comparables o mayores que las previamente reportadas en estudios de entrenamiento aeróbico de una duración similar.
Agradecimientos
Los autores agradecen a John Moroz por la asistencia técnica y a nuestros sujetos por su tiempo y esfuerzo.
Dirección para el Envío de Correspondencia
M. J. Gibala, Exercise Metabolism Research Group, Dept. of Kinesiology, McMaster Univ., Hamilton, Ontario, Canada L8S 4K1 (correo electrónico: gibalam@mcmaster.ca).
Referencias
1. Barnett C., Carey M., Proietto J., Cerin E., Febbraio M. A., and Jenkins D (2004). Muscle metabolism during sprint exercise in man: influence of sprint training. J Sci Med Sport 7: 314322
2. Bogdanis G. C., Nevill M. E., Boobis L. H., and Lakomy H. K (1996). Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J Appl Physiol 80: 876884
3. Bonen A., McCullagh K. J., Putman C. T., Hultman E., Jones N. L., and Heigenhauser G. J (1998). Short-term training increases human muscle MCT1 and femoral venous lactate in relation to muscle lactate. Am J Physiol Endocrinol Metab 274: E102E107
4. Bradford M. M (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248254
5. Carter S. L., Rennie C. D., Hamilton S. J., and Tarnopolsky M. A (2001). Changes in skeletal muscle in males and females following endurance training. Can J Physiol Pharmacol 79: 386392
6. Chesley A., Heigenhauser G. J., and Spriet L. L (1996). Regulation of muscle glycogen phosphorylase activity following short-term endurance training. Am J Physiol Endocrinol Metab 270: E328E335
7. Dawson B., Fitzsimons M., Green S., Goodman C., Carey M., and Cole K (1998). Changes in performance, muscle metabolites, enzymes and fibre types after short sprint training. Eur J Appl Physiol 78: 163169
8. Fluck M. and Hoppeler H (2003). Molecular basis of skeletal muscle plasticity-from gene to form and function. Rev Physiol Biochem Pharmacol 146: 159216
9. Gorostiaga E. M., Walter C. B., Foster C., and Hickson R. C (1991). Uniqueness of interval and continuous training at the same maintained exercise intensity. Eur J Appl Physiol 63: 101107
10. Green H. J., Barr D. J., Fowles J. R., Sandiford S. D., and Ouyang J (2004). Malleability of human skeletal muscle Na+-K+-ATPase pump with short-term training. J Appl Physiol 97: 143148
11. Green H. J., Cadefau J., Cusso R., Ball-Burnett M., and Jamieson G (1995). Metabolic adaptations to short-term training are expressed early in submaximal exercise. Can J Physiol Pharmacol 73: 474482
12. Green H., Grant S., Bombardier E., and Ranney D (1999). Initial aerobic power does not alter muscle metabolic adaptations to short-term training. Am J Physiol Endocrinol Metab 277: E39E48
13. Harmer A. R., McKenna M. J., Sutton J. R., Snow R. J., Ruell P. A., Booth J., Thompson M. W., Mackay N. A., Stathis C. G., Crameri R. M., Carey M. F., and Eager D. M (2000). Skeletal muscle metabolic and ionic adaptations during intense exercise following sprint training in humans. J Appl Physiol 89: 17931803
14. Harris R. C., Hultman E., and Nordesjo L. O (1974). Glycogen, glycolytic intermediates and high-energy phosphates determined in biopsy samples of musculus quadriceps femoris of man at rest: methods and variance of values. Scand J Clin Lab Invest 33: 109120
15. Henriksson J., Chi M. M., Hintz C. S., Young D. A., Kaiser K. K., Salmons S., and Lowry O. H (1986). Chronic stimulation of mammalian muscle: changes in enzymes of six metabolic pathways. Am J Physiol Cell Physiol 251: C614C632
16. Henriksson J. and Reitman J. S (1976). Quantitative measures of enzyme activities in type I and type II muscle fibres of man after training. Acta Physiol Scand 97: 392397
17. Holloszy J. O. and Coyle E. F (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol 56: 831838
18. Jacobs I., Esbjornsson M., Sylven C., Holm I., and Jansson E (1987). Sprint training effects on muscle myoglobin, enzymes, fiber types and blood lactate. Med Sci Sports Exerc 19: 368374
19. Juel C., Klarskov C., Nielsen J. J., Krustrup P., Mohr M., and Bangsbo J (2004). Effect of high-intensity intermittent training on lactate and H+ release from human skeletal muscle. Am J Physiol Endocrinol Metab 286: E245E251
20. Krustrup P., Hellsten Y., and Bangsbo J (2004). Intense interval training enhances human skeletal muscle oxygen uptake in the initial phase of dynamic exercise at high but not at low intensities. J Physiol 559: 335345
21. Leek B. T., Mudaliar S. R., Henry R., Mathieu-Costello O., and Richardson R. S (2001). Effect of acute exercise on citrate synthase activity in untrained and trained human skeletal muscle. Am J Physiol Regul Integr Comp Physiol 280: R441R447
22. Leppik J. A., Aughey R. J., Medved I., Fairweather I., Carey M. F., and McKenna M. J (2004). Prolonged exercise to fatigue in humans impairs skeletal muscle Na+-K+-ATPase activity, sarcoplasmic reticulum Ca2+ release, and Ca2+ uptake. J Appl Physiol 97: 14141423
23. Linossier M. T., Denis C., Dormois D., Geyssant A., and Lacour J. R (1993). Ergometric and metabolic adaptation to a 5-s sprint training programme. Eur J Appl Physiol 67: 408414
24. MacDougall J. D., Hicks A. L., MacDonald J. R., McKelvie R. S., Green H. J., and Smith K. M (1998). Muscle performance and enzymatic adaptations to sprint interval training. J Appl Physiol 84: 21382142
25. McKenna M. J., Heigenhauser G. J., McKelvie R. S., Obminski G., MacDougall J. D., and Jones N. L (1997). Enhanced pulmonary and active skeletal muscle gas exchange during intense exercise after sprint training in men. J Physiol 501: 703716
26. Nevill M. E., Boobis L. H., Brooks S., and Williams C (1989). Effect of training on muscle metabolism during treadmill sprinting. J Appl Physiol 67: 23762382
27. Ortenblad N., Lunde P. K., Levin K., Andersen J. L., and Pedersen P. K (2000). Enhanced sarcoplasmic reticulum Ca2+ release following intermittent sprint training. Am J Physiol Regul Integr Comp Physiol 279: R152R160
28. Parolin M. L., Chesley A., Matsos M. P., Spriet L. L., Jones N. L., and Heigenhauser G. J (1999). Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am J Physiol Endocrinol Metab 277: E890E900
29. Parra J., Cadefau J. A., Rodas G., Amigo N., and Cusso R (2000). The distribution of rest periods affects performance and adaptations of energy metabolism induced by high-intensity training in human muscle. Acta Physiol Scand 169: 157165
30. Passoneau J. V. and Lowry O. H. Enzymatic (1993). Analysis: A Practical Guide. Totowa, NJ: Humana
31. Putman C. T., Jones N. L., Hultman E., Hollidge-Horvat M. G., Bonen A., McConachie D. R., and Heigenhauser G. J (1998). Effects of short-term submaximal training in humans on muscle metabolism in exercise. Am J Physiol Endocrinol Metab 275: E132E139
32. Rodas G., Ventura J. L., Cadefau J. A., Cusso R., and Parra J (2000). A short training programme for the rapid improvement of both aerobic and anaerobic metabolism. Eur J Appl Physiol 82: 480486
33. Ross A. and Leveritt M (2001). Long-term metabolic and skeletal muscle adaptations to short-sprint training: implications for sprint training and tapering. Sports Med 15: 10631082
34. Sale D. G (1991). Testing strength and power. In: Physiological Testing of the High-Performance Athlete (2nd ed.), edited by MacDougall JD, Wenger HA, and Green HA. Champaign, IL: Human Kinetics, p. 7182
35. Sandiford S. D., Green H. J., Duhamel T. A., Perco J. G., Schertzer J. D., and Ouyang J (2004). Inactivation of human muscle Na+-K+-ATPase in vitro during prolonged exercise is increased with hypoxia. J Appl Physiol 96: 17671775
36. Shoemaker J. K., Phillips S. M., Green H. J., and Hughson R. L (1996). Faster femoral artery blood velocity kinetics at the onset of exercise following short-term training. Cardiovasc Res 31: 278286
37. Spina R. J., Chi M. M., Hopkins M. G., Nemeth P. M., Lowry O. H., and Holloszy J. O (1996). Mitochondrial enzymes increase in muscle in response to 710 days of cycle exercise. J Appl Physiol 80: 22502254
38. Tonkonogi M., Harris B., and Sahlin K (1997). Increased activity of citrate synthase in human skeletal muscle after a single bout of prolonged exercise. Acta Physiol Scand 161: 435436
39. Trump M. E., Heigenhauser G. J., Putman C. T., and Spriet L. L (1996). Importance of muscle phosphocreatine during intermittent maximal cycling. J Appl Physiol 80: 15741580
40. Tupling A. R (2004). The sarcoplasmic reticulum in muscle fatigue and disease: role of the sarco(endo)plasmic reticulum Ca2+-ATPase. Can J Appl Physiol 29: 308329