
Efectos del Entrenamiento de Fuerza de Alta Intensidad sobre los Patrones de Respuesta Hormonal en Hombres Jóvenes y Ancianos
Effects of Heavy-Resistance Training on Hormonal Response Patterns in Younger Vs. Older Men
Keijo Häkkinen3, William J Kraemer1, Robert U Newton4, Bradley C Nindl2, Jeff S Volek2, Matthew McCormick2, Lincoln A Gotshalk2, Scott E Gordon2, Steven J Fleck6 y Wayne W Campbell5
1Human Perfomance Laboratory, Ball State University, Muncie, Indiana, U.S.A.
2Center for Sports Medicine/Noll Physiological Research Center, Pennsylvania State University, University Park, Pennsylvania 16802.
3Department of Biology of Physical Activity, University of Jyväskilä, Finland and Research Unit for Sport and Physical Fitness, Jyväskilä, Finland.
4School of Exercise Science and Sport Management, Southern Cross University, Lismore, New South Wales, Australia.
5Nutrition, Metabolism, and Exercise Laboratory, Donald W. Reynolds Department of Geriatrics, University of Arkansas for Medical Sciences, North Little Rock, Arkansas 72114-1706.
6Department of Sport Science, Colorado College, Colorado Springs, Colorado 80913.
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 3, Número 167 del año 2022.
Publicado 28 de julio de 2003
Resumen
Palabras clave: endocrino, envejecimiento, sarcopenia, fuerza, músculo, andropausia, somatopausia, factores de crecimiento
Abstract
Keywords: endocrine, aging, sarcopenia, strength, muscle, andropause, somatopause, growth factors
INTRODUCCION
El envejecimiento normal está asociado con la reducción en la masa magra, particularmente en la masa muscular (i.e., sarcopenia) con la subsecuente disminución en la fuerza muscular y en las capacidades funcionales (18-20, 44, 60, 62). Con la edad, las concentraciones circulantes de hormonas anabólicas y de factores de crecimiento [e.g., hormona del crecimiento (GH), testosterona y factores de crecimiento de tipo insulínico (IGF)-I] también se reducen (i.e., somatopausia y andropausia) (4, 11, 16, 22, 33, 42, 46, 56, 61, 62). A lo largo de los últimos 15 años, el entrenamiento de la fuerza ha constituido la principal intervención utilizada para compensar los efectos del envejecimiento a través de la minimización de la reducción de la fuerza, la masa magra y las capacidades funcionales relacionadas con el envejecimiento (2, 14, 18, 20, 54). Campbell y cols. (2) demostraron que el entrenamiento de la fuerza puede mejorar positivamente la retención de nitrógeno y el metabolismo corporal total de proteínas musculares en hombres ancianos. La mejora del turnover o recambio proteico favorece el crecimiento del tejido muscular que esta bajo la regulación homeostática interactiva del sistema endócrino (5, 6, 17, 31, 32, 50, 70). Los cambios hormonales producidos por el entrenamiento de pesas pueden constituir un estímulo importante para los hombres ancianos, contribuyendo a la prevención de la sarcopenia, la perdida de fuerza y la perdida de las capacidades funcionales. Sin embargo, los datos acerca de los efectos del entrenamiento de la fuerza sobre los cambios en los patrones de respuesta hormonal en hombres ancianos son limitados (5, 18, 54).
Se ha mostrado que, en hombres jóvenes, el entrenamiento de la fuerza de alta
intensidad constituye un potente estímulo para el incremento agudo de las
hormonas circulantes (5, 23, 24, 30, 36-38, 40, 41, 59). En contraste, en
hombres ancianos, el entrenamiento de la fuerza de alta intensidad, no ha
mostrado provocar una respuesta hormonal de igual magnitud (i.e., >60 años de
edad) (5, 23, 54, 59). La importancia del nivel de hormonas circulantes reside
en el hecho de que se piensa que las menores cantidades de hormonas anabólicas
circulantes influencian la reducción en la masa muscular relacionada al
envejecimiento y la capacidad de fuerza asociada, en la sexta década de vida (4,
17, 61, 62). Debido a que se ha observado que las alteraciones en las
concentraciones hormonales en hombres jóvenes se producen en la fase temprana
del entrenamiento (67), la cuestión que emerge es si en los hombres ancianos
pueden ocurrir mecanismos hormonales similares en la fase temprana de un
programa de entrenamiento de la fuerza. Dichos cambios, sin embargo, no han sido
frecuentemente observados (5, 18, 20, 54). Esto podría deberse al tipo de
programa de entrenamiento de la fuerza utilizado, ya que algunos protocolos no
parecen ser efectivos para estimular las respuestas hormonales debido a la baja
intensidad, la poca masa muscular involucrada y/o los largos períodos de reposo
entre las series y los ejercicios (16, 24, 34, 36, 38). Además solo algunos
pocos estudios han presentado datos del conjunto entero de hormonas anabólicas y
catabólicas y de las respuestas agudas y crónicas al ejercicio de sobrecarga y
al entrenamiento (23, 54).
Si un programa de entrenamiento de la fuerza apropiado (e.g., un programa
periodizado) puede aumentar los niveles de reposo y/o las respuestas inducidas
por el ejercicio, entonces el mismo podría de valor terapéutico para establecer
cambios endocrinos en hombres ancianos. El principal propósito de esta
investigación fue examinar el patrón de los cambios en las concentraciones
hormonales de reposo e inducidas por el ejercicio en hombres jóvenes y ancianos,
antes y después de un programa periodizado de entrenamiento de la fuerza
diseñado para incrementar la fuerza y el tamaño muscular.
METODOS
Sujetos
Ocho hombres jóvenes (30 años de edad [30Y]) y nueve hombres ancianos (62 años de edad [62Y]) fueron voluntarios para participar en esta investigación, la cual fue aprobada por el Comité de Revisión Institucional para la Utilización de Sujetos Humanos de la Universidad Estatal de Pennsylvania. Los riesgos y beneficios del estudio fueron completamente explicados a todos los sujetos, y subsiguientemente se obtuvo su consentimiento escrito. Previamente a su inclusión en el estudio todos los sujetos fueron sometidos una revisión médica y debían ser aprobados por el médico como saludables (i.e., libres de cualquier problema ortopédico, endócrino o cualquier otro problema médico). Todos los sujetos eran físicamente activos, pero no habían estado involucrados en programas estructurados de entrenamiento de la fuerza. Se utilizaron cuestionarios de actividad física los cuales revelaron que todos los sujetos estaban involucrados en la práctica de deportes recreacionales y de jogging. Las características de los sujetos antes del entrenamiento eran las siguientes: para el grupo 30Y la edad media fue 29.8±5.3 años, la masa corporal fue de 90.0±12.8kg, la altura fue de 177.2±5.3cm y el porcentaje de grasa corporal fue de 18.3±4.6%; para el grupo 62Y la edad media fue de 62±3.2 años, la masa corporal fue de 84.3±13.4kg, la altura fue de 177.0±7.3cm y el porcentaje de grasa corporal fue de 20.4±4.6%. De la información recolectada, se observó que las características físicas y los antecedentes de actividad física de estos sujetos eran esencialmente similares, siendo la edad la única diferencia significativa entre los dos grupos (p≤0.05).
Composición Corporal
La composición corporal fue valorada mediante la medición de pliegues cutáneos. Todas las mediciones antropométricas fueron obtenidas por el mismo investigador en el lado derecho de los sujetos. Los espesores de los pliegues cutáneos se obtuvieron con un calibre para pliegues cutáneos Harpenden (Country Technology, Gays Mills, WI; 10g/mm de presión constante) y se midieron los pliegues del pecho, axilar medial, abdominal, suprailíaco, subescapular, tricipital y muslo siguiendo procedimientos previamente descriptos (45). Las mediciones fueron repetidas hasta que se obtuvieran dos mediciones dentro de 1mm, utilizando la media de estas dos mediciones. Para estimar la densidad corporal se utilizó una ecuación que utiliza los valores de los siete pliegues cutáneos (29), y subsiguientemente se calculó el porcentaje de grasa corporal utilizando la ecuación de Siri (66). No hubo diferencias significativas en el porcentaje de grasa entre los grupos.
Área de Sección Transversal Muscular
El área de sección cruzada muscular del muslo (MCSA) de la pierna dominante fue valorada antes y después del entrenamiento de la fuerza por medio de un imán superconductor de 0.5-T (Picker International, Highland Heights, OH) de imágenes de resonancia magnética (MRI) equipado con un programa MR6B. Las imágenes fueron obtenidas por medio de la alteración del spin-lattice o tiempo de relajación longitudinal (T1). Las imágenes T1 fueron obtenidas utilizando un tiempo de repetición de 500ms, un tiempo de eco de 13ms, un radio de frecuencia de 90º, y una potencia de absorción de 0.028W/kg.
La MCSA fue analizada a partir de las imágenes de MRI por medio de la técnica de gradiente de eco, la cual permite la mayor delineación y distinción entre los músculos. Una vez que el sujeto estaba posicionado dentro del imán, la pierna dominante era apoyada por debajo de la rodilla para que estuviera paralela a la camilla del MRI, y los pies eran sujetados con una correa para evitar la rotación. Luego de esto se obtenía una imagen sagital del muslo. Sobre la imagen sagital se colocó una cuadrícula de 15 secciones y se obtuvieron imágenes transaxiales. Se obtuvieron 15 imágenes transaxiales del muslo de 1 cm de espesor, equidistantes entre la base de la cabeza femoral y la mitad de la articulación de la rodilla. Todas las imágenes MRI fueron entonces introducidas en una computadora Macintosh para el cálculo de las MCSA individuales y totales utilizando un programa modificado del Instituto Nacional de Salud (NIH). Para calcular la MCSA, se utilizó la sección 8 (siendo la sección 1 la base de la cabeza femoral). El área de sección transversal del tejido se obtuvo por medio del programa NIH 1.55.20A para el análisis de imágenes a través de la contabilización de pixeles. El mismo investigador realizó todas las mediciones de áreas, y el coeficiente de correlación intraclase para la confiabilidad test-retest fue r=0.99 (21).
Evaluación de una Repetición Máxima
La fuerza máxima fue valorada por medio del test de 1 repetición máxima (RM, i.e., la mayor carga que pudiera ser levantada correctamente una vez) en sentadilla solamente concéntrica a partir de un ángulo de la rodilla de 90º (37, 73) con la utilización de un dispositivo Sistema de Potencia Pliométrico (Norsearch, Lismore, Australia). La entrada en calor consistió de una serie de 5-10 repeticiones al 40-60% del máximo estimado. Una vez realizada la entrada en calor los sujetos descansaron por ~1min y realizaron algunos estiramientos suaves. Luego de esto se realizaron de tres a cinco repeticiones con el 60-80% del máximo estimado. Para determinar la 1RM se realizaron tres a cuatro intentos con 3-5min de pausa entre los levantamientos. Para que cada prueba de 1RM se considerara exitosa se requirió que estas fueran realizadas con la técnica apropiada y en el rango completo de movimiento. Durante la evaluación de la 1RM no se produjeron lesiones. El coeficiente de confiabilidad test-retest para este test fue R=0.97.
Test de Ejercicio Agudo de Fuerza de Alta Intensidad
Cada sujeto se familiarizó con el protocolo experimental del test de ejercicio agudo de fuerza de alta intensidad (AHRET) y realizó el mismo antes y después de finalizar las 10 semanas de entrenamiento. El AHRET consistió de cuatro series de 10RM del ejercicio de sentadilla con 90s de pausa entre las series. Si debido a la aparición de fatiga en alguna de las series, el sujeto no podía realizar las 10 repeticiones, se realizaba un ajuste de la carga (e.g., reducción de la carga) para permitir que el sujeto pudiera completar las 10 repeticiones en la siguiente serie. Las sentadillas fueron realizadas en el dispositivo Sistema de Potencia Pliométrico, como se describió previamente (73).
Recolección y Análisis de las Muestras de Sangre
Una semana antes del entrenamiento (pre) y al finalizar el mismo (post) se obtuvieron muestras de sangre antes de la realización del AHRET, inmediatamente después (IP), y a los 5, 15 y 30min post ejercicio utilizando una cánula que se mantuvo abierta mediante una traba salina. Además, utilizando técnicas similares, se recolectaron muestras de sangre en la semana –3 (3 semanas antes del comienzo del entrenamiento) y en las semanas 0, 3, 6 y 10 durante el entrenamiento. Las muestras de sangre obtenidas antes del entrenamiento fueron utilizadas como muestras de control para realizar comparaciones, y para ilustrar con mayor claridad los datos obtenidos, en los RESULTADOS solo se describen los valores de la semana 0. Las muestras de sangre recolectadas en las semanas –3 y 0 fueron indistinguibles (p>0.05). Para limitar la influencia de cualquier variación diurna (1, 11, 28, 64), durante la investigación, las muestras de sangre fueron obtenidas en diferentes horas del día entre los sujetos, pero a la misma hora del día para cada sujeto. Las muestras de sangre se recolectaron luego del ayuno nocturno, y además se monitoreo y registró la ingesta dietaria a través de los distintos puntos del tiempo. Durante la recolección de muestras de reposo, en la semana 0, en dos de los sujetos del grupo 30Y y en uno de los sujetos del grupo 62Y las muestras de sangre se extrajeron fuera de la ventana de 90 minutos (dentro de las 2.5hs). Las muestras de sangre fueron centrifugadas a 1500g y a –4ºC durante 15 minutos. Todas las muestras de suero y de plasma fueron colocadas en tubos con conservantes y guardadas a –84ºC hasta la realización de los análisis. El suero se obtuvo para los análisis de testosterona total (TT), testosterona libre (FT), cortisol, GH y lactato, mientras que el plasma heparinizado se obtuvo para el análisis de ACTH. Los valores de reposo para las concentraciones séricas de IGF-I y de proteína-3 ligadora de IGF (IGFBP-3) se determinaron solamente pre y post-entrenamiento.
El hematócrito se determinó por triplicado utilizando la técnica estándar de microcapilaridad. La Hb se determinó calorimétricamente por duplicado utilizando el método de cianometahemoglobina (Sigma Chemical, St. Louis, MO). Todas las hormonas fueron analizadas por duplicado utilizando las diferentes técnicas de RIA. Para eliminar la varianza interanálisis, todas las muestras fueron analizadas en una misma tanda de análisis, y todas las varianzas intraensayo fueron <5%. La testosterona serica (TT), la FT y el cortisol fueron analizados por medio del radioinmunoensayo 125I de anticuerpo único (fase sólida), mientras que para la ACTH y la GH sérica se utilizó el radioinmunoensayo de doble anticuerpo (Diagnostic Products, Los Angeles, CA). La concentración de IGF-I se analizó por medio de radioinmunoensayo 125I de fase líquida y doble anticuerpo con la extracción preliminar de una columna de octadecil silil silica para separar los IGF-I de sus proteínas de unión (INCSTAR, Stillwater, MN). Las IGFBP-3 se midieron utilizando el ensayo inmunoradiométrico 125I de dos sitios (Diagnostic Systems Laboratory, Webster, TX). La sensibilidad de los anticuerpos fue de 0.14nmol/l para la TT, 0.52pmol/l para la FT, 0.19pmol/l para la ACTH, 5.5nmol/l para el cortisol, <2nmol/l para los IGF-I, 0.5ng/ml para las IGFBP-3 y 0.9 μg/l para la GH. Para determinar los valores de radioinmunoactividad se utilizó un contador gamma modelo LKB 1272 Clini y un sistema de reducción de datos en línea (Pharmacia LKB Nuclear, Gaithersburg, MD). Las concentraciones de lactato corporal total se midieron en duplicado utilizando un analizador de lactato (Sport Lactate Analyzer 1500, Yellow Springs Instruments, Yellow Springs, OH). Los cambios en el volumen plasmático se calcularon utilizando los valores del hematócrito y de la Hb, utilizando los métodos descritos por Dill y Costill (7). Las concentraciones hormonales no fueron corregidas por los cambios en el volumen plasmático. Los cambios en el volumen plasmático en este estudio entre las condiciones pre y el post-entrenamiento AHRET (IP) fueron -11.4±6.2 y -14.0±5.6% pre-entrenamiento y -10.4±5.2 y -12.0±4.6% post-entrenamiento para los grupos 30Y y 62Y, respectivamente. No se observaron diferencias significativas entre los grupos o con el entrenamiento.
Programa Periodizado de Sobrecarga para el Entrenamiento de la Fuerza/Potencia
Todos los sujetos se familiarizaron con todos los protocolos de ejercicio utilizados en este estudio. Además, todas las sesiones de ejercicio fueron supervisadas individualmente. El programa periodizado de entrenamiento de la fuerza utilizó un modelo de periodización no lineal y consistió de múltiples ejercicios y múltiples series realizados tres veces por semana durante 10 semanas (12, 13). Las sesiones diarias fueron alternadas variando la carga (intensidad) y el volumen (series x repeticiones x carga) a lo largo de la semana. Los días lunes se realizaron series de 3-5RM con 2-3 minutos de pausa entre las series. Los miércoles, se realizaron series de 8-10RM con 1 minuto de pausa entre las series. Finalmente, los viernes la carga fue de ~12-15RM, sin embargo, se realizaron solamente 6-8 repeticiones, con la intensión de realizar estos ejercicios con una mayor producción de potencia o de manera explosiva. Se permitieron 1-2min de pausa entre las series. El programa consistió de los siguientes ejercicios: sentadilla, extensiones de rodilla, extensiones de la espalda, dorsales en polea, flexiones de rodilla, elevaciones de pantorrilla, press de banca, remo sentado, press militar y curl de bíceps.
Análisis Estadísticos
Los datos fueron analizados utilizando análisis de varianza ANOVA de dos vías para mediciones repetidas, para los análisis post hoc se utilizó el test de Fisher de la menor diferencia significativa para hallar diferencias en las medias apareadas. La fortaleza estadística de los cálculos para este estudio osciló entre 0.78 y 0.80. Se estableció el nivel de significancia a p≤0.05.
RESULTADOS
Fuerza e Hipertrofia
El grupo 30Y tuvo una mayor 1RM en sentadilla y una MCSA media del muslo mayor que el grupo 62Y, tanto antes como después del programa de entrenamiento. Luego de 10 semanas de entrenamiento periodizado de la fuerza, la fuerza en 1RM en sentadilla se incrementó desde 139±22 hasta 163±23kg en el grupo 30Y y desde 102±34 hasta 113±37kg en el grupo 62Y. La MCSA se incrementó desde 186±16 hasta 204±18cm2 y desde 159±22 hasta 169±26cm2 luego del entrenamiento periodizado de la fuerza en los grupos 30Y y 62Y, respectivamente. No se observaron cambios significativos en la masa corporal o en el porcentaje de grasa corporal luego del entrenamiento en ninguno de los dos grupos. La Figura 1 muestra el porcentaje de incremento en la fuerza en 1RM en el ejercicio de sentadilla y la MCSA. Tanto el grupo 30Y como el grupo 62Y experimentaron incrementos porcentuales similares para la 1RM en sentadilla (~15%), pero el grupo 30Y tuvo una mayor cantidad de hipertrofia en la MCSA que el grupo 62Y (10.1±3.7 y 5.9±2.9% para el grupo 30Y y 62Y, respectivamente). Asimismo, se observó un incremento del 8.4±4.6% (p≤0.05) en la MCSA calculado en forma separada para el grupo muscular de los isquiotibiales en el grupo 30Y, que fue mayor (p≤0.5) que el incremento de 3.6±3.3% registrado para el grupo 62Y. Mientras que los incrementos de 12.2±3.6 y 8.4±4.6% para el cuadriceps no difirieron significativamente entre los grupos.
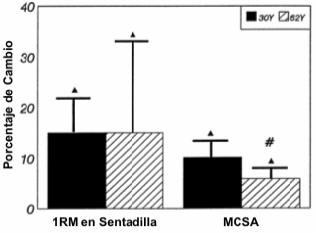
Figura 1. Porcentaje de cambio (media±DE) entre las condiciones pre hasta
el post-entrenamiento para el grupo de hombres jóvenes (30Y) y para el grupo de
hombres ancianos (62Y) para las variables 1RM y área de sección transversal
total del muslo (MCSA). ▲ Significativamente diferente (p≤0.05) con respecto al
valor pre-entrenamiento correspondiente; # diferencia estadísticamente
significativa (p≤0.05) entre los grupos 30Y y 62Y.
Concentraciones Hormonales en Reposo
Las Figuras 2-4 describen los cambios en las concentraciones hormonales en reposo para la TT, FT, ACTH y cortisol en las semanas 0, 3, 6 y 10 y para el IGF-I e IGFBP-3, tanto en las condiciones pre como post-entrenamiento. Para la TT (Figura 2A), no se observaron diferencias significativas tanto para la edad como para el entrenamiento. Sin embargo, los efectos de interacción indicaron mayores concentraciones de TT en las semanas 6 y 10 en el grupo 30Y en comparación con en grupo 62Y. Se observaron efectos de interacción significativos de la edad para la respuesta de la FT (Figura 2B). En la semana 0, no hubo diferencias significativas en la FT entre los grupos 30Y y 62Y, pero aparecieron diferencias significativas en la semana 3 que persistieron hasta el final del programa de entrenamiento. Además, para el grupo 30Y, las concentraciones séricas de FT se elevaron en la semana 10 en comparación con la semana 0. Para el grupo de hombres de 62Y, la FT se mantuvo sin cambios a lo largo del programa de entrenamiento. No se observaron efectos de la edad, el entrenamiento o de interacción para la edad o el entrenamiento sobre las concentraciones de ACTH (Figura 3A). Para el cortisol (Figura 3B) se observaron efectos de interacción que indicaban una menor concentración en la semana 3 y en la semana 10 para el grupo 62Y.
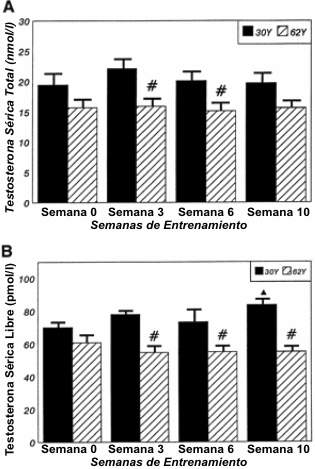
Figura 2. Concentraciones en reposo (media±DE) para la testosterona total
(A) y para la testosterona libre (B) durante las 10 semanas de entrenamiento
periodizado de la fuerza y potencia. ▲ Significativamente diferente (p≤0.05) con
respecto valor correspondiente en la semana 0; # diferencia estadísticamente
significativa (p≤0.05) entre los grupos 30Y y 62Y.
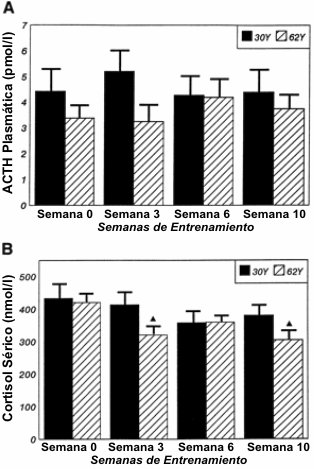
Figura 3. Concentraciones en reposo (media±DE) para la ACTH (A) y para el
cortisol (B) durante las 10 semanas de entrenamiento periodizado de la fuerza y
potencia. ▲ Significativamente diferente (p≤0.05) con respecto valor
correspondiente en la semana 0.
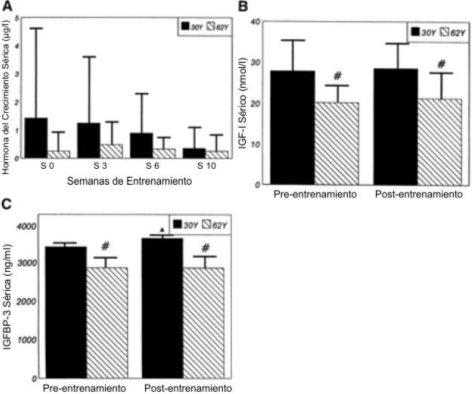
Figura 4. Concentraciones en reposo (media±DE) para la hormona del
crecimiento (A), para el factor de crecimiento de tipo insulínico tipo I (IGF-I;
B), y para la proteína ligadora de IGF-I (IGFBP-3; C) durante las 10 semanas de
entrenamiento periodizado de la fuerza y potencia. ▲ Significativamente
diferente (p≤0.05) con respecto valor correspondiente en la semana 0; #
diferencia estadísticamente significativa (p≤0.05) entre los grupos 30Y y 62Y.
Para la GH (Figura 4A), no se observaron efectos de la edad, entrenamiento o de interacción. Las Figuras 4B y 4C muestran las mediciones pre y post-entrenamiento de las concentraciones séricas de IGF-I y de IGFBP-3 séricas. La concentración sérica de IGF-I (Figura 4B) fue mayor en el grupo 30Y que en el grupo 62Y tanto en la condición pre como post-entrenamiento. Además, desde el pre al post-entrenamiento no se produjeron cambios en la concentración de IGF-I en ninguno de los dos grupos. Las concentraciones de IGFBP-3 fueron mayores en el grupo 30Y que en el grupo 62Y, tanto antes como después del entrenamiento (Figura 4C). Sin embargo, el grupo 30Y mostró un incremento en la concentración de IGFBP-3 luego de las 10 semanas de entrenamiento de la fuerza, lo que no ocurrió en el grupo 62Y.
Respuestas Hormonales al AHRET
Las Figuras 5-7 describen las respuestas hormonales y del lactato luego de la realización del AHRET antes y después de las 10 semanas de entrenamiento. Para la TT (Figura 5A) se observaron efectos significativos debido a la edad. Las medias para los efectos principales debido a la edad fueron 20.3 y 15.7nmol/l para los grupos 30Y y 62Y, respectivamente. Antes del entrenamiento, se observaron elevaciones por encima de los valores basales, inmediatamente después del AHRET y a los 5 y 15 minutos de la recuperación en el grupo 30Y e inmediatamente después del AHRET y a los 15 minutos de la recuperación para el grupo 62Y. A los 30 minutos de recuperación, las concentraciones de TT para ambos grupos fueron similares a los valores pre-ejercicio. El grupo 30Y mostró elevaciones, inmediatamente después y 5 minutos después de la realización del AHRET. El grupo 62Y mostró elevaciones en la TT inmediatamente después de la realización de AHRET y a los 5 minutos de recuperación. Los efectos de interacción para el grupo 62Y también indicaron que los valores de la TT post-entrenamiento a los 5 y 30 minutos de la recuperación fueron mayores que los valores pre-entrenamiento correspondientes.
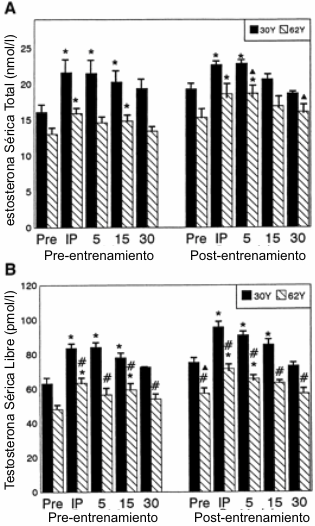
Figura 5. Respuesta de la testosterona total (A) y de la testosterona
libre (B) luego de la realización del test de ejercicio agudo de fuerza de alta
intensidad (AHRET), antes y después de 10 semanas de entrenamiento periodizado
de la fuerza y potencia en los grupos 30Y y 62Y. * Significativamente diferente
(p≤0.05) con respecto al valor correspondiente pre-ejercicio; # diferencia
estadísticamente significativa (p≤0.05) entre los grupos 30Y y 62Y; ▲
significativamente diferente (p≤0.05) con respecto al valor correspondiente en
la condición pre-entrenamiento. Pre, pre ejercicio. IP, inmediatamente después
del ejercicio; 5, 15 y 30: 5, 15 y 30 minutos después del ejercicio.
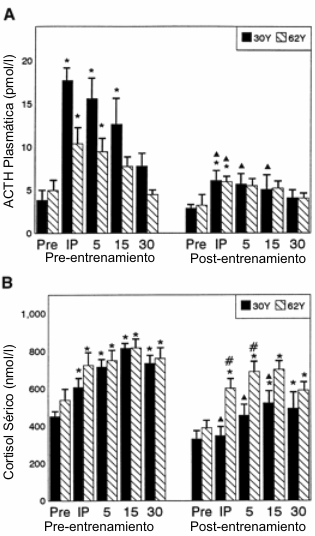
Figura 6. Respuestas (media±DE) de la ACTH (A) y del cortisol (B) luego
de la realización del AHRET, antes y después de 10 semanas de entrenamiento
periodizado de la fuerza y potencia en los grupos 30Y y 62Y. *
Significativamente diferente (p≤0.05) con respecto al valor correspondiente pre-ejercicio;
# diferencia estadísticamente significativa (p≤0.05) entre los grupos 30Y y 62Y;
▲ significativamente diferente (p≤0.05) con respecto al valor correspondiente en
la condición pre-entrenamiento.
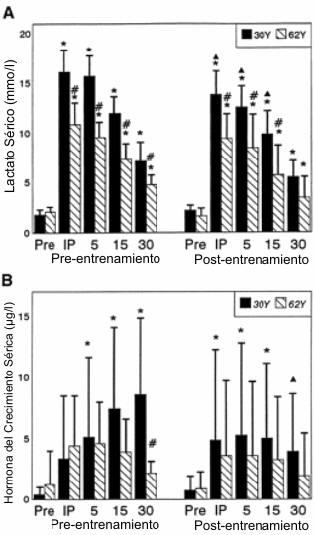
Figura 7. Respuestas (media±DE) del lactato (A) y la hormona del
crecimiento (B) luego de la realización del AHRET, antes y después de 10 semanas
de entrenamiento periodizado de la fuerza y potencia en los grupos 30Y y 62Y. *
Significativamente diferente (p≤0.05) con respecto al valor correspondiente pre-ejercicio;
# diferencia estadísticamente significativa (p≤0.05) entre los grupos 30Y y 62Y;
▲ significativamente diferente (p≤0.05) con respecto al valor correspondiente en
la condición pre-entrenamiento.
Para la FT (Figura 5B), se hallaron efectos significativos para la edad, tiempo de recuperación y entrenamiento. Las medias para los efectos principales de la edad fueron 80 y 60pmol/l para el grupo 30Y y 62Y, respectivamente. Las medias para los efectos principales debidos al entrenamiento fueron 66 y 74pmol/l, antes y después del entrenamiento, respectivamente. Para los efectos principales del tiempo de recuperación, se observaron elevaciones por encima de los valores basales inmediatamente después del AHRET y a los 5 y 15 minutos de recuperación, pero no a los 30 minutos. Los efectos de interacción revelaron diferencias entre los grupos 30Y y 62Y en todos los puntos del tiempo, excepto para el valor pre-ejercicio antes del entrenamiento. También se observaron elevaciones por encima de los valores pre-ejercicio inmediatamente después y a los 5 y 15 minutos posteriores a la realización del AHRET en el grupo 30Y, tanto en las condiciones pre como post-entrenamiento. En el grupo 62Y, se hallaron valores de FT mayores a los correspondientes en la condición pre-ejercicio inmediatamente después y a los 15min post AHRET, pre-entrenamiento, e inmediatamente después y a los 5min post AHRER, post-entrenamiento.
En comparación con los valores pre-entrenamiento para la ACTH (Figura 6A), después del entrenamiento se observaron reducciones significativas luego de la realización del AHRET. En el grupo 30Y, antes del entrenamiento la ACTH se elevó inmediatamente después del ejercicio y a los 5 y 15min; sin embrago, después del entrenamiento, las elevaciones fueron evidentes solamente inmediatamente post-ejercicio. En el grupo 62Y, antes del entrenamiento, la ACTH se elevó inmediatamente después del ejercicio y a los 5 minutos, pero solamente inmediatamente post-ejercicio, luego del entrenamiento. Además, en el grupo 30Y los valores post-entrenamiento obtenidos inmediatamente después del ejercicio y a los 5 y 15 minutos fueron menores que los valores correspondientes a los obtenidos antes del entrenamiento. En el grupo 62Y los valores obtenidos inmediatamente después del ejercicio fueron menores post-entrenamiento que pre-entrenamiento. El cortisol se elevó por encima de los valores pre AHRET en todos los puntos del tiempo en ambos grupos antes del entrenamiento (Figura 6B). Después del entrenamiento, en el grupo 62Y el cortisol se elevó por encima de los valores basales en todos los puntos del tiempo. En el grupo 30Y, el cortisol se elevó por encima de los valores pre-ejercicio a los 15 y 30min post-ejercicio. Los valores obtenidos para este grupo inmediatamente después del ejercicio y a los 15 y 30min de la recuperación fueron menores que los valores correspondientes antes del entrenamiento. Se observaron diferencias entre los grupos en los valores obtenidos inmediatamente después del ejercicio y a los 5min de recuperación, después del entrenamiento.
El lactato se elevó significativamente por encima de los valores basales después de la realización del AHRET en ambos grupos, tanto antes como después del programa de entrenamiento (Figura 7A). Los valores de lactato fueron mayores en el grupo 30Y en comparación con el grupo 62Y en cualquiera de las mediciones, excepto a los 30 minutos post AHRET después del entrenamiento. En el grupo 30Y los valores de lactato post-entrenamiento obtenidos inmediatamente después del ejercicio y a los 5 y 15min post AHRET, fueron menores que los correspondientes valores antes del entrenamiento. En el grupo 62Y no se observaron elevaciones de la GH (Figura 7B) por encima de los valores pre AHRET, tanto antes como después entrenamiento. En el grupo 30Y se observaron valores elevados a los 5, 15 y 30 minutos de recuperación, antes del entrenamiento. Después del entrenamiento, en el grupo 30Y, los valores obtenidos inmediatamente post-ejercicio y a los 5 y 15 minutos de recuperación fueron mayores que los valores basales pre-ejercicio. Para el grupo 30Y el valor obtenido a los 30min de recuperación fue menor que el valor correspondiente antes del entrenamiento.
DISCUSION
Las respuestas hormonales en este estudio reflejan los cambios en la adaptación en un grupo de hombres de 60 años de edad muy activos y con buena aptitud física, quienes fueron capaces de realizar un programa muy intenso de entrenamiento de la fuerza corporal total. La eficacia y las adaptaciones asociadas a dicho programa de entrenamiento de la fuerza en sujetos de 60 años de edad con menor aptitud física requieren estudios adicionales. Sin embargo, se han utilizado programas de entrenamiento de la fuerza para ancianos, los cuales pueden requerir de una progresión más apropiada para asegurar la tolerancia a protocolos más avanzados (13).
Las reducciones en las concentraciones de hormonas anabólicas (e.g., testosterona, GH, e IGF) con la edad pueden influenciar las reducciones en el tamaño muscular y en la fuerza, observadas con el envejecimiento (32, 50, 63). La restauración de la función de las glándulas endócrinas con el ejercicio constituye una hipótesis atractiva, lo cual puede ayudar a aliviar las reducciones en la masa de tejido muscular y en el fuerza relacionadas con el envejecimiento. En este estudio hemos examinado las adaptaciones en la fase temprana de las hormonas circulantes a un programa periodizado de entrenamiento de la fuerza de alta intensidad que tenía como objetivo el incremento del tamaño muscular y de la fuerza. Aunque la fase temprana del entrenamiento de la fuerza esta caracterizada por considerables adaptaciones neurales (e.g., incremento en la activación de los músculos agonistas), las adaptaciones intrínsecas al músculo pueden también tener lugar en la fase temprana del entrenamiento (67). En un estudio previo con hombres jóvenes, las adaptaciones en la fase temprana incluyeron transformaciones en los tipos de fibras musculares e incrementos en la fuerza que fueron asociados con incrementos tempranos en la concentración de testosterona sérica y con reducciones en la concentración sérica de cortisol (67). El presente estudio tuvo la característica única de que utilizó un programa periodizado de entrenamiento de la fuerza de alta intensidad, el cual fue efectivo para provocar un incremento en la masa muscular y en la fuerza en solo 10 semanas. Uno de los principales hallazgos del presente estudio fue que los hombres jóvenes mostraron un incremento significativamente mayor en el tamaño total del muslo, aunque las ganancias relativas, al menos en la fuerza de los extensores de la rodilla, no fueron diferentes entre los grupos de jóvenes y de ancianos. La capacidad de los hombres jóvenes para provocar una respuesta hipertrófica relativamente mayor en es este estudio parece estar asociada a las diferencias en los patrones de adaptación hormonal tanto en reposo como inducidos por el ejercicio. El otro hallazgo principal de este estudio fue que el sistema endócrino mostró tener una “plasticidad” para los cambios adaptativos en hombres ancianos y jóvenes en la fase temprana del programa periodizado de entrenamiento de la fuerza de alta intensidad.
En los hombres, la testosterona es una hormona anabólica potente que media la acumulación de proteínas y también mejora la función neural (6, 51, 70). En general, la concentración de testosterona inducida por el ejercicio (e.g., efecto principal para la TT [20.3 vs. 15.7nmol/l] y la FT [80 vs. 60pmol/l]) fueron significativamente mayores en el grupo 30Y en comparación con el grupo 62Y. Además, los análisis del área bajo la curva revelaron una mayor magnitud de incremento en la FT inducida por el ejercicio en los hombres jóvenes como respuesta al estimulo del entrenamiento de la fuerza. Estos datos respaldan el concepto de “andropausia”, el cual está caracterizado por reducciones en el número de células de Leydig en los testículos, reducciones en la capacidad secretoria, y una reducción en la secreción de gonadotropinas, tanto en reposo como durante la estimulación (69-71). Staron y cols. (67) demostraron que las concentraciones de reposo de TT en hombres con una edad media de 23.5 años, se incrementaban por encima de los valores basales luego de solo 4 semanas de entrenamiento. Sin embargo, otros estudios no han mostrado un incremento relacionado con el entrenamiento en la concentración de reposo de testosterona. Esto indica que, en hombres jóvenes, la testosterona puede exhibir un patrón de respuesta hormonal homeostática dinámica al entrenamiento de la fuerza. Sin embargo, en un estudio realizado por Nicklas y cols. (54) no se observaron alteraciones en las concentraciones de reposo de TT a lo largo de un programa progresivo de entrenamiento de 16 semanas en hombres con un promedio de edad de 60 años. Häkkinen y Pakarinen (25) observaron que en hombres de 70 años de edad tampoco se producía dicha respuesta. A pesar de la falta de cambios en las concentraciones hormonales, estos autores reportaron relaciones significativas entre las concentraciones de testosterona y los cambios en la fuerza con el entrenamiento en hombres de 70 años (r=0.61). En el presente estudio, aunque no se observaron cambios en las concentraciones de testosterona en reposo durante las 10 semanas de entrenamiento en el grupo de hombres de 62Y, este grupo demostró una mejora en la capacidad para estimular la liberación de TT luego de la realización de ejercicios de sobrecarga. Este hallazgo es único y puede estar relacionado al programa periodizado de entrenamiento, sin embargo, las razones exactas y el/los mecanismo/s fisiológico/s que median esta adaptación permanecen bajo especulación.
El/los mecanismo/s que median dicha adaptación en las concentraciones séricas de testosterona inducidas por el ejercicio en hombres ancianos podría deberse a incrementos en la liberación pulsátil o en la producción de la hormona luteinizante (LH) (34, 46, 51, 69, 71). Sin emabrgo, Lu y cols. (47) reportaron que el incremento en la concentración de testosterona en ratas macho durante el ejercicio fue, al menos parcialmente, el resultado de un mecanismo estimulatorio del ejercicio (LH-independiente), en donde el lactato influencia la secreción de la testosterona por medio del incremento en la producción de AMPc testicular. Evidencia reciente también implica al oxido nítrico (a través de mecanismos vasodilatadores) y a patrones del flujo sanguíneo que impactan sobre la liberación de testosterona a nivel de los testículos (49). El impacto de estos cambios inducidos por el ejercicio en los hombres ancianos es poco claro, pero podría haber contribuido al incremento en el tamaño muscular y al incremento de la fuerza observado en este estudio. Uno podría especular que en los hombres ancianos dichos incrementos adaptativos en la testosterona podrían haber contribuido en mayor grado a los cambios relativos similares en al fuerza, a través de la influencia de la testosterona sobre los mecanismos neurales (e.g., incremento en la síntesis de neurotransmisores) (36, 51). Se ha demostrado que los factores neurales son muy importantes en la mediación de los incrementos de la fuerza con el entrenamiento a corto plazo en hombres ancianos (19, 52, 53). De hecho, los cambios relativos similares en la fuerza de los extensores de la rodilla junto con una menor magnitud de incremento en el área de sección transversal total del muslo (y también del cuadriceps femoral, aunque no estadísticamente diferente del grupo 30Y) en hombres ancianos parecen respaldar la teoría de que los factores neurales juegan un papel importante para explicar la producción de fuerza en 1RM (19, 52, 53, 58). Más específicamente, ambos grupos demostraron un menor incremento en el tamaño de los isquiotibiales que el observado en el cuadriceps femoral, y el grupo 62Y mostró un menor incremento inducido por el entrenamiento en los músculos isquiotibiales que el respectivo incremento registrado en el grupo 30Y. Consistentemente con los datos presentes, se ha demostrado en individuos ancianos (65) menores incrementos en el tamaño del grupo muscular de los isquiotibiales que los observados en el cuadriceps femoral. Debido a que la fuerza de los flexores de la rodilla no fue registrada, en este estudio no fue posible evaluar la contribución de los factores neurales en el desarrollo de la fuerza de los isquiotibiales entre los grupos 30Y y 62Y.
Las concentraciones de reposo de FT fueron mayores en los hombres jóvenes a lo largo de la mayor parte del período de entrenamiento, y nuevamente los hombres jóvenes mostraron la capacidad de provocar una mayor respuesta de la FT inducida por el ejercicio en comparación con los hombres ancianos. Häkkinen y Pakarinen (25) previamente demostraron la importancia de la FT biológicamente activa para la entrenabilidad de los hombres jóvenes. La testosterona circula en sangre unida principalmente a una globulina ligadora de hormonas sexuales o a albúmina. Aunque se piensa que la testosterona libre (se considera FT tanto a la hormona que circula libre como a la testosterona que circula unida a la albúmina) es regulada por la TT, el complejo de testosterona unida, con un mayor peso molecular, ha mostrado se incapaz de atravesar el endotelio capilar y de penetrar la membrana plasmática para interactuar con los elementos regulatorios del núcleo (27, 36, 48, 57). Por lo tanto la mayor concentración de FT en la sangre parece indicar una mejora del “nivel de bioactividad” en los hombres jóvenes (51, 55, 56). Sobre todo, un mayor ambiente androgénico en los hombres jóvenes en reposo y luego de las sesiones de ejercicio parece respaldar la mayor magnitud total del incremento observado en el tamaño muscular del muslo. Además, estos datos nos dan pruebas potenciales de las diferencias relacionadas con la edad en el desarrollo del tejido muscular con programas de entrenamiento a corto plazo, aunque estas diferencias pueden no ser del mismo grado para todos los músculos o grupos musculares. De esta manera, parece haber una ventaja relacionada con la edad en el mecanismo mediado por la FT para responder a un programa de entrenamiento de la fuerza en la fase temprana del mismo.
Otro hallazgo importante en este estudio fue que la cantidad de cortisol producida en reposo fue reducida y la respuesta al estrés producido por el entrenamiento de la fuerza fue menor en los hombres ancianos. Las reducciones en las concentraciones de de cortisol en reposo a través del programa de entrenamiento en los hombres ancianos, sin cambios significativos en las concentraciones de ACTH, indica que los receptores de ACTH en la glándula adrenal pueden haber sufrido una “reducción” (27, 35). Con el entrenamiento, se han observado reducciones en la respuesta del cortisol al estrés producido por el ejercicio de sobrecarga solamente en hombres jóvenes. En un estudio previo, Staron y cols. (67) mostraron una respuesta similar en hombres jóvenes. Estos cambios en la respuesta del cortisol inducida por el ejercicio luego de la realización de ejercicio son aparentemente mediados por la reducción en la respuesta de la ACTH al estrés provocado por el ejercicio de sobrecarga. Se piensa que las reducciones en el cortisol proveen un mecanismo posible por el cual se aumenta la acumulación de proteínas, a través de la reducción de la degradación de las fibras musculares tipo I (8, 36). Los hombres ancianos pueden también depender de la hipertrofia de las fibras musculares tipo I para provocar una mayor hipertrofia muscular total debido a la pérdida de fibras musculares tipo II con el proceso de envejecimiento (43). Por lo tanto la importancia de estos cambios en el cortisol no puede ser minimizada, especialmente en hombres ancianos, como mecanismo relacionado a la hipertrofia del tejido muscular y a la capacidad para producir fuerza. Colectivamente, estos datos indican que los hombres ancianos pueden provocar una reducción en las respuestas de las hormonas catabólicas, lo que resulta en un ambiente anabólico más favorable para reducir la degradación de proteínas o para incrementar la síntesis de las mismas.
En este estudio no se observaron cambios significativos en la concentración de reposo de la GH a lo largo del programa de entrenamiento, tanto en los sujetos jóvenes como en los ancianos. Estos resultados están respaldados por hallazgos similares publicados en la literatura (5, 15, 25, 26, 54, 67). La respuesta del lactato al ejercicio fue menor en los hombres ancianos que en los jóvenes con la utilización de un estrés de ejercicio relativamente similar. Los hombres jóvenes también mostraron una respuesta del lactato ligeramente reducida a la misma intensidad relativa de ejercicio luego del entrenamiento, y esto puede explicar parcialmente porque algunos de los valores post-ejercicio de la GH fueron bajos, ya que se ha demostrado que los mecanismos ácido-base están relacionados a la liberación de GH (15). Aunque en los estudios de entrenamiento de la fuerza con hombres ancianos comúnmente no se han observado cambios en la adaptación de la GH, las respuestas de la GH luego de la realización de un ejercicio continuo posterior al entrenamiento de la resistencia son reducidas (72). Debido a la naturaleza pulsátil de la GH, las mediciones aisladas en reposo deben ser interpretadas con precaución, especialmente en hombres jóvenes, quienes muestran una secreción pulsátil mucho mayor (11, 71). Nuestros datos son consistentes con los hallazgos de otros estudios de entrenamiento de la fuerza y GH en hombres jóvenes y ancianos (5, 25, 54, 67). En general, los incrementos significativos en el patrón de liberación de GH inducido por el ejercicio luego del entrenamiento de la fuerza se han observado solamente en el grupo 30Y. Esto coincide con estudios previos en los cuales el ejercicio no produjo una respuesta de la GH en hombres ancianos (5, 54) y en los cuales la respuesta de la GH se redujo luego del entrenamiento de la fuerza en ciertos, pero no en todos, los puntos del tiempo de la recuperación (63). Estos resultados pueden deberse en parte a diferencias en la edad de los sujetos o a diferencias en los programas de entrenamiento (i.e., series totales, repeticiones, periodos de recuperación y ejercicios realizados). Esta es solamente una comprensión rudimentaria de los efectos del ejercicio y el envejecimiento sobre los mecanismos fisiológicos que subyacen a la “somatopausia” (i.e., la disminución del sistema GH-IGF-I) (4, 61). Debido a que se piensa que este eje endocrino es de gran importancia para mantener la integridad del sistema musculoesquelético, es conceptualmente pragmático sugerir que los regímenes de ejercicio deberían tener como objetivo la estimulación de este eje. La GH también exhibe una gran heterogeneidad molecular, y el radioinmunoensayo estándar se enfoca en la variante de 22-kDa (3, 10, 68). Esto es un punto especialmente importante para las futuras investigaciones, cuando se considere que las especies de GH de mayor peso molecular podrían poseer una mayor actividad biológica y que la falta de cambios o las reducciones en la inmunorreactividad de la GH pueden no representar el esquema completo de respuestas de adaptación de la GH al entrenamiento de la fuerza (3, 10, 15, 68).
Similarmente a otros estudios, el IGF-I fue más bajo en el grupo 62Y que en el grupo 30Y (4, 54). Las diez semanas de entrenamiento no afectaron esta diferencia relacionada a la edad, y por lo tanto el entrenamiento a corto plazo no parece ser efectivo para alterar la concentración de IGF-I (54). Sin embargo, un hallazgo particular de este estudio fue que el entrenamiento de la fuerza produjo un incremento significativo en la concentración de IGFBP-3 en el grupo de hombres jóvenes. Además, la concentración de IGFBP-3 fue mayor que en el grupo de ancianos tanto antes como después del entrenamiento. Se cree que el IGF-I y la IGFBP-3 se liberan en el hígado, aunque pocos estudios han examinado los efectos del entrenamiento crónico de la fuerza sobre los componentes del sistema IGF. En estudios de entrenamiento de la resistencia, las IGFBP-3 han mostrado ser independientes de las respuestas de los IGF-I y que potencialmente tienen su propia actividad biológica a nivel celular (32). Recientemente, Elikiam y cols. (9) mostraron que las concentraciones musculares de IGF-I pueden incrementarse con el entrenamiento de la resistencia, a pesar de la falta de cambios en el RNAm IGF-I muscular o en la concentración sérica de IGF-I. La falta de cambio en la concentración sérica de IGF-I con el entrenamiento puede sugerir que los IGF-I en la circulación pueden no ser un marcador significativo de la actividad implícita del sistema GH-IGF-I. El IGF-I pueden operar en el músculo más que de manera autocrina/paracrina, o el IGF-I liberado por el incremento en la secreción desde el hígado puede ser rápidamente secuestrado por el tejido en un esfuerzo por mantener el equilibrio homeostático del mismo en la circulación sistémica. El IGF-I provoca una inhibición negativa de la secreción de GH al nivel del hipotálamo y de la pituitaria. El secuestro de IGF-I por el tejido podría servir para aumentar la acción mitógenica, y al mismo tiempo proteger al sistema de la reducción en la liberación hipotalámica y pituitaria de GH.
En resumen, este estudio ha demostrado las diferencias en las concentraciones hormonales entre hombres con promedios de edad de 30 y 62 años. Sin embargo, el entrenamiento periodizado de la fuerza es muy efectivo para producir ganancias iniciales en el tamaño muscular y en la fuerza. Estas adaptaciones están asociadas más probablemente con las adaptaciones neurales tempranas de los músculos entrenados y también con los cambios en el sistema endócrino tanto en hombres ancianos como jóvenes. La incapacidad de emplear mecanismos hormonales similares en respuesta al ejercicio de sobrecarga de alta intensidad indica que la plasticidad del sistema endocrino en los hombres ancianos se ha alterado o desmejorado. Sin embrago, este estudio fue el primero en demostrar que los hombres ancianos pueden tener adaptaciones fisiológicas en el sistema endócrino con el entrenamiento de la fuerza. Si períodos más largos de entrenamiento periodizado de la fuerza en ancianos pueden continuar produciendo adaptaciones hormonales adicionales que estén asociadas con posteriores cambios en el tamaño muscular, la fuerza y las capacidades funcionales, nos deja una hipótesis provocativa para un futuro estudio.
Agradecimientos
Queremos agradecer a la Dra. Arja Häkkinen, al Dr. J. Michael Lynch, a los estudiantes graduados y a los miembros del laboratorio por su ayuda y respaldo a este proyecto. También queremos agradecer al muy dedicado grupo de sujetos que participaron en la investigación e hicieron que este proyecto fuera posible.
Nota al Pie
Queremos agradecer el respaldo a este proyecto por un subsidio otorgado por el Ministerio de Educación de Finlandia.
Referencias
1. Brandenberger, G., and M. Follenius (1975). Influence of timing and intensity of muscular exercise on temporal patterns of plasma cortisol levels. J. Clin. Endocrinol. Metab. 40: 845-849
2. Campbell, W. W., M. C. Crim, V. R. Young, L. J. Joseph, and W. J. Evans (1995). Effects of resistance training and dietary protein intake on protein metabolism in older adults. Am. J. Physiol. 268 (Endocrinol. Metab. 31): E1143-E1153
3. Charrier, J., and J. Martal (1988). Growth hormones. 1. Polymorphism. Reprod. Nutr. Dev. 28: 857-887
4. Copeland, K. C., R. B. Colletti, J. T. Devlin, and T. L. McAuliffe (1990). The relationship between insulin-like growth factor-I and aging. Metabolism 39: 584-587
5. Craig, B. W., R. Brown, and J. Everhart (1989). Effects of progressive resistance training on growth hormone and testosterone levels on young and elderly subjects. Mech. Ageing Dev. 49: 159-169
6. 6. Crist, D. M., G. T. Peake, R. B. Loftfield, J. C. Kraner, and P. A. Egan (1991). Supplemental growth hormone alters body composition, muscle protein metabolism and serum lipids in fit adults: characterization of dose-dependent and response-recovery effects. Mech Ageing Dev, 58: 191-205
7. Dill, D. B., and D. L. Costill (1974). Calculation of percentage changes in volume of blood, plasma, and red cells in dehydration. J Appl Physiol, 37: 247-248
8. Doerr, P., and K. Pirke (1976). Cortisol-induced suppression of plasma testosterone in normal adult males. J Clin Endocrinol Metab, 43: 622-628
9. Elikiam, A., M. Moromisato, D. Moromisato, J. A. Brasel, C. Roberts, and D. M. Cooper (1997). Increase in muscle IGF-I protein but not IGF-I mRNA after 5 days of endurance training in young rats. J Appl Physiol, 42: 1557-1561
10. Ellis, S., M. A. Vodian, and R. E. Grindeland (1978). Studies on the bioassayable growth hormone-like activity of plasma. Recent Prog Horm Res, 34: 213-238
11. Finklestein, J. W., H. P. Roffwarg, R. M. Boyar, J. Kream, and L. Hellman (1972). Age-related change in the twenty-four hour spontaneous secretion of growth hormone. J Clin Endocrinol Metab, 35: 665-670
12. Fleck, S. J., and W. J. Kraemer (1996). Periodization Breakthrough. Ronkonkoma, NY: Advanced Research
13. Fleck, S. J., and W. J. Kraemer (1997). Designing Resistance Training Programs (2nd ed.). Champaign, IL: Human Kinetics
14. Frontera, W. R., C. N. Meridith, K. P. O'Reilly, H. G. Knuttgen, and W. J. Evans (1983). Strength training in older men: skeletal muscle hypertrophy and improved function. J Appl Physiol, 64: 1038-1044
15. Gordon, S. E., W. J. Kraemer, N. H. Vos, J. M. Lynch, and H. G. Knuttgen (1994). Effect of acid-base balance on the growth hormone response to acute high-intensity cycle exercise. J Appl Physiol, 76: 821-829
16. Gray, A., H. A. Feldman, J. B. McKinlay, and C. Longcope (1991). Age, disease, and changing sex hormone levels in middle-aged men: results of the Massachusetts male aging study. J Clin Endocrinol Metab, 73: 1016-1025
17. Griggs, R. C., W. Kingston, R. F. Jozefowicz, B. E. Herr, G. Forbes, and D. Halliday (1989). Effects of testosterone on muscle mass and muscle protein synthesis. J Appl Physiol, 66: 498-503
18. Hakkinen, K., R. U. Newton, S. E. Gordon, M. R. McCormick, J. Volek, B. C. Nindl, L. A. Gotshalk, W. Campbell, W. J. Evans, A. Hakkinen, B. Humphries, and W. J. Kraemer (1998). Changes in muscle morphology, electromyographic activity, and force production during periodized strength training in young and old men. J Gerontol A Biol Sci Med Sci, 53: 415-423
19. Hickson, R. C., and J. R. Marone (1993). Exercise and inhibition of glucocorticoid-induced muscle atrophy. In: Exercise and Sport Sciences Reviews. Baltimore, MD: Williams & Wilkins, vol. 21, p. 135-167
20. Horrocks, P. M., A. F. Jones, W. A. Ratcliffe, G. Holder, A. White, R. Holder, J. G. Ratcliffe, and D. R. London (1990). Patterns of ACTH and cortisol pulsatility over twenty-four hours in normal males and females. Clin Endocrinol (Oxf.), 32: 127-134
21. Jackson, A. S., and M. L. Pollock. (1978). Generalized equations for predicting body density of men. Br J Nutr, 40: 497-504
22. Jensen, J., H. Oftebro, B. Breigan, A. Johnson, K. Ohlin, H. D. Meen, S. B. Stromme, and H. A. Dahl (1991). Comparison of changes in testosterone concentrations after strength and endurance exercise in well trained men. Eur J Appl Physiol, 63: 467-471
23. Johannsson, G., G. Grimby, K. S. Sunnerhagen, and B. Bengstom (1997). Two years of growth hormone (GH) treatment increase isometric and isokinetic muscle strength in GH-deficient adults. J Clin Endocrinol Metab 82: 2877-2884
24. Jones, J. I., and D. R. Clemmons (1995). Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev, 16: 3-34
25. Kern, W., C. Dodt, J. Born, and H. L. Fehm (1996). Changes in cortisol and growth hormone secretion during nocturnal sleep in the course of aging. J Gerontol A Biol.Sci Med Sci, 51: M3-M9
26. Kraemer, W. J (1988). Endocrine responses to resistance exercise. Med Sci Sports Exerc, 20: S152-S157
27. Kraemer, W. J., S. J. Fleck, R. Callister, M. Shealy, G. A. Dudley, C. M. Maresh, L. Marchitelli, C. Cruthirds, T. Murray, and J. E. Falkel (1989). Training responses of plasma -endorphin, adrenocorticotropin, and cortisol. Med Sci Sports Exerc, 21: 146-153
28. Kraemer, W. J., S. J. Fleck, and W. J. Evans. (1996). Strength and power training: physiological mechanisms of adaptation. In: Exercise and Sport Sciences Reviews, edited by J. O. Holloszy. Baltimore, MD: Williams & Wilkins, vol. 24, p. 363-397
29. Kraemer, W. J., S. E. Gordon, S. J. Fleck, L. J. Marchitelli, R. Mello, J. E. Dziados, K. Friedl, E. A. Harman, C. Maresh, and A. C. Fry (1991). Endogenous anabolic hormonal and growth factor response to heavy resistance exercise in males and females. Int J Sports Med, 12: 228-235
30. Kraemer, W. J., L. Marchitelli, S. E. Gordon, E. A. Harman, J. E. Dziados, R. Mello, P. Frykman, D. McCurry, and S. J. Fleck (1990). Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol, 69: 1442-1450
31. Kraemer, W. J., B. J. Noble, M. J. Clark, and B. W. Culver (1987). Physiologic responses to heavy-resistance exercise with very short rest periods. Int J Sports Med, 8: 247-252
32. Kraemer, W. J., J. F. Patton, S. E. Gordon, E. A. Harman, M. R. Deschenes, K. Reynolds, R. U. Newton, T. N. Triplett, and J. E. Dziados (1995). Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J Appl Physiol, 83: 976-989
33. Kraemer, W. J., J. F. Patton, H. G. Knuttgen, L. J. Marchitelli, C. Cruthirds, A. Damokosh, E. A. Harman, P. Frykman, and J. E. Dziados (1989). Hypothalamic-pituitary-adrenal responses to short duration high-intensity cycle exercise. J Appl Physiol, 66: 161-166
34. Lambers, S. W. J., A. W. van den Beld, and A. J. van der Lely (1997). The endocrinology of aging. Science, 278: 419-424
35. Larsson, L. (1982). Physical training effects on muscle morphology in sedentary males at different ages. Med Sci Sports Exerc, 14: 203-206
36. Larsson, L., and J. Karlsson (1978). Isometric and dynamic endurance as a function of age and skeletal muscle characteristics. Acta Physiol Scand, 104: 129-136
37. Lohman, T. G., A. F. Roche, and R. Martorell (1988). Anthropometric Standardization Reference Manual. Champaign, IL: Human Kinetics
38. Longcope, C., S. R. W. Goldfield, D. J. Brambilla, and J. McKinlay (1990). Androgens, estrogens, and sex hormone-binding globulin in middle-aged men. J Clin Endocrinol Metab, 71: 1442-1446
39. Lu, S. S., C. P. Pang, Y. F. Tung, S. W. Huang, Y. H. Chen, H. C. Shih, S. C. Tsai, C. C. Lu, S. W. Wang, J. J. Chen, E. J. Chien, C. H. Chien, and P. S. Wang (1997). Lactate and the effects of exercise on testosterone secretion: evidence for the involvement of a cAMP mediated mechanism. Med Sci Sports Exerc, 29: 1048-1054
40. Manni, A., W. M. Pardridge, W. Cefalu, B. C. Nisula, C. W. Bardin, S. J. Santer, and R. J. Santen (1985). Bioavailability of albumin-bound testosterone. J Clin Endocrinol Metab, 61: 705-710
41. Meskaitis, V. J., F. S. Harman, J. S. Volek, B. C. Nindl, W. J. Kraemer, D. Weinstock, and D. R. Deaver (1997). Effects of exercise on testosterone and nitric oxide production in the rat testis (Abstract). J Androl, Jan-Feb, Suppl: P-37
42. Marcus, R., G. Butterfield, L. Holloway, L. Gilliland, D. J. Baylink, R. L. Hintz, and B. M. Sherman (1990). Effects of short term administration of recombinant human growth hormone to elderly people. J Clin Endocrinol Metab, 70: 517-527
43. Mooradian, A. D., J. E. Morley, and S. G. Korenman (1987). Biological actions of androgens. Endocr Rev, 8: 1-17
44. Morotani, T (1992). Time course of adaptations during strength and power training. In: The Encyclopedia of Sports Medicine: Strength and Power in Sport, edited by P. V. Komi. London: Blackwell Scientific, p. 266-278
45. Moritani, T., and H. A. DeVries (1980). Neural factors versus hypertrophy in the time course of muscle strength gains. Am J Phys Med 82: 521-524
46. Nicklas, B. J., A. J. Ryan, M. M. Treuth, S. M. Harman, M. R. Blackman, B. F. Hurley, and M. A. Rogers (1995). Testosterone, growth hormone and IGF-1 responses to acute and chronic resistive exercise in men aged 55-70 years. Int J Sports Med, 16: 445-450
47. Ninkin, H. R., and J. H. Calkins (1986). Decreased bioavailability of testosterone in aging normal and impotent men. J Clin Endocrinol Metab 63: 1418-1420
48. Partridge, W. M (1987). Plasma protein-mediated transport of steroid and thyroid hormones. Am J Physiol 252 (Endocrinol. Metab. 15): E157-E164
49. Ploutz, L. L., P. A. Tesch, R. L. Biro, and G. A Dudley (1994). Effect of resistance training on muscle use during exercise. J Appl Physiol 76: 1675-1681
50. Pyka, G., D. R. Taaffe, and R. Marcus (1994). Effect of a sustained program of resistance training on the acute growth hormone response to resistance exercise in older adults. Horm Metab Res, 26: 330-333
51. Rogers, M. A., and W. J. Evans (1993). Changes in skeletal muscle with aging: effects of exercise training. In: Exerc and Sport Sci Rev. Balt, MD: Williams & Wilkins, vol. 21, p. 65-102
52. Rosen, C. J., and C. Conover (1997). Growth hormone/insulin-like growth factor-I axis in aging: a summary of a National Institute on Aging-sponsored symposium. J Clin. Endocrinol Metab, 82: 3919-3922
53. Rudman, D (1985). Growth hormone, body composition and aging. J Am Geriatr Soc, 33: 800-807
54. Rudman, D., M. H. Kutner, C. M. Rogers, M. F. Lubin, G. A. Fleming, and R. P. Gain (1981). Impaired growth hormone secretion in the adult population: relation to age and adiposity. J Clin Invest, 67: 1361-1369
55. Sherman, B., C. Wysham, and J. B. Pfohl (1985). Age-related changes in the circadian rhythm of plasma cortisol in man. J Clin Endocrinol Metab, 61: 439-443
56. Siri, W. E (1961). Body composition from fluid spaces and density: analysis of methods. In: Tech for Meas Body Comp, edi by J. Brozek, and A. Henschel. Wash, DC: Natl Acad Sci, p. 223-244
57. Staron, R. S., D. L. Karapondo, W. J. Kraemer, A. C. Fry, S. E. Gordon, J. E. Falkel, F. C. Hagerman, and R. S. Hikida (1994). Skeletal muscle adaptations during early phase of heavy-resistance training in men and women. J Appl Physiol, 76: 1247-1255
58. Stolar, M. W., and G. Baumann (1986). Big growth hormone forms in human plasma: immunochemical evidence for their pituitary origin. Metabolism, 35: 75-77
59. Tenover, J. L (1997). Testosterone and the aging male. J.Androl, 19: 103-106
60. Tenover, J. S (1994). Androgen administration to aging men. Clin Androl, 23: 877-893
61. Vermeulen, A., R. Rubens, and L. Verdonck (1972). Testosterone secretion and metabolism in male senescence (Abstract). J Clin Endocrinol, 34: 730
62. Weltman, A., J. Y. Weltman, C. J. Womack, S. E. Davis, J. L. Blumer, G. A. Gaesser, and M. L. Hartman (1997). Exercise training decreases the growth hormone (GH) response to acute constant-load exercise. . Med Sci Sports Exerc, 29: 669-676
63. Wilson, G. J., R. U. Newton, A. J. Murphy, and B. J. Humphries (1993). The optimal training load for the development of dynamic athletic performance. Med Sci Sports Exerc, 25: 1279-1286