
Fundamentos neurofisiológicos del aprendizaje de la técnica deportiva
Publicado 13 de julio de 2015, 7:02

Formación de la memoria como punto central del aprendizaje motor
La memoria es indispensable para todos los procesos de aprendizaje y de adaptación, pues toda modificación del comportamiento se basa en una evaluación comparativa, una valoración y un proyecto nuevo.
La formación de la memoria se basa en mecanismos de adaptación neurofisiológicos complicados y no aclarados definitivamente hasta la fecha.
Según el estado actual de los conocimientos (cf. Matthies,1973, 531 y 1979, 179; Ott, 1977, 104; Kokonen, 1979, 50; Lössner y cols., 1979, 125; Rahmann 1979, 107; Voronin/Danilova 1979, 121 s.; Wenzel/Kammerer/Frotscher 1979, 361; Kugler 1981, 5 s.), la formación de la memoria, y en consecuencia también el aprendizaje motor, se puede explicar por procesos metabólicos neuronales, que originan en último término alteraciones duraderas de las membranas sinápticas, y por tanto una diferente permeabilidad para los diferentes aflujos de excitación (informaciones codificadas).
Así pues, podemos definir el aprendizaje del movimiento y de la técnica como un condicionamiento de enlaces sinápticos, que produce un nuevo reticulado de los sistemas neuronales específicos del movimiento.
La diversidad y el volumen de los procesos metabólicos neuronales en el transcurso del aprendizaje son inabarcables: en un segundo de actividad mental normal –un aspecto del aprendizaje motor– se metabolizan en cada célula del cerebro unas 15.000 moléculas de proteína (Kugler, 1981, 5).
La figura 1 ofrece un resumen de los procesos metabólicos y estructurales que tienen lugar durante el aprendizaje motor.
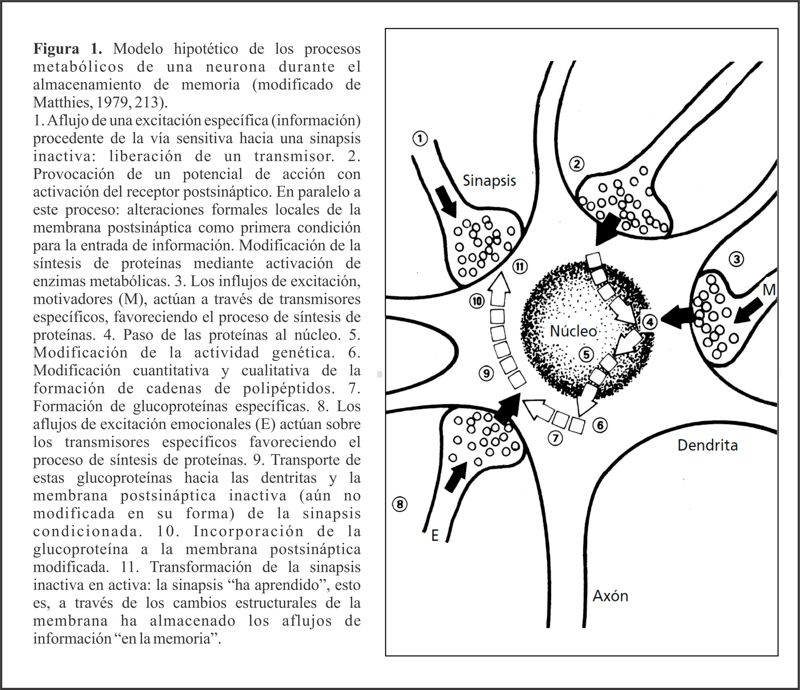
| El aprendizaje del movimiento y de la técnica se basa en procesos bioquímicos que discurren en estructuras anatómicas jerárquicamente ordenadas (ver Tabla 1), y que se organizan, mediante alteraciones sinápticas, de forma superpuesta en un reticulado específico de sistemas neuronales. |
![]()
La existencia de productos de síntesis que condicionan la transmisión de información ha podido demostrarse de forma convincente con el aislamiento de péptidos responsables de alteraciones del comportamiento; pensemos en la “escotofobina”, aislada y producida de forma sintética por Ungar (1973, 317). Desde entonces cientos de experimentos han conseguido transmitir informaciones delimitadas y aprendidas con ayuda de las sustancias condicionantes específicas (cf. Gay/Raphaelson, 1967; McConnell/Malin, 1973, 343; Fjerdingstad, 1973, 430; Domagk/Schonne/Thines, 1973, 419).
Memoria inmediata, memoria a corto plazo y memoria a largo plazo
La existencia de la memoria inmediata –llamada también memoria de segundos– se atribuye sobre todo a procesos bioeléctricos que tienen su origen en los neurotransmisores (cf. Guttmann/Matwyshin/Weiler, 1973, 397; Huston/ Mueller, 1979, 176; Kugler, 1981, 8).
El correlato de la memoria a corto plazo paree ser el proceso siguiente, originado a su vez por los procesos bioeléctricos. Según Sinz (1977, 204), después de 500 ms de la entrada de una información específica del aprendizaje se producen ya reacciones neuroquímicas, que ponen en marcha la síntesis de moléculas proteicas en elementos estructurales pericarionales (situados alrededor del núcleo).
La estructura química de la proteína formada depende probablemente de la combinación de procesos electrofisiológicos en la membrana y de los neurotransmisores y moduladores participantes en el proceso.
La síntesis de las moléculas proteicas necesita un tiempo determinado –entre unos minutos y unos días–, hasta que, a partir de la llamada iniciación, se llega a la formación completa de una molécula proteica (cf. Kugler, 1981, 8).
Finalmente, la memoria a largo plazo aparece una vez concluido el transporte de la molécula proteica sintetizada hacia lugares determinados de la membrana y su alojamiento en la estructura de lípidos de dicha membrana.
Así pues, las memorias inmediata, a corto y a largo plazo se basan en una secuencia de procesos aislados (fig. 2) que se condicionan mutuamente. El conjunto del proceso se puede modificar en diferentes etapas de su desarrollo mediante influjos de estimulación o de inhibición (fig. 1).
| Todos los enfoques parciales muestran que los mecanismos de formación de la memoria y las fases de la memoria basadas en éstos sólo se pueden entender como resultado conjunto de la interacción de mecanismos reguladores moleculocelulares y de acontecimientos en el nivel de la red neuronal (Ott, 1977, 104). |

Reforzadores de memoria
Determinadas neurohormonas influyen de manera específica sobre el proceso de aprendizaje y formación de la memoria (cf. Stark/Ott/Matthies, 1979, 315). Su efecto se plasma en el refuerzo o el mantenimiento de los procesos de la memoria. Constituyen la base de la memoria a corto plazo y modulan los procesos siguientes, que permiten la memoria a largo plazo.
Junto a los refuerzos “positivos” distinguimos también otros “negativos”, cuyo efecto es un empeoramiento del efecto del aprendizaje (cf. Huston/Muller, 1979, 175).
Entre los refuerzos positivos encontramos una serie de péptidos con un efecto específico en el cerebro, procedentes de los lóbulos anterior (p. ej., ACTH), intermedio (p. ej., alfa-MSH) o posterior (p. ej., vasopresina) de la hipófisis. Todos estos péptidos elevan la resistencia ante el olvido de materias de aprendizaje; se diferencian sólo en la duración de su efecto; algunos actúan durante horas (p. ej., ACTH), días (p. ej., DS1–15) o semanas (p. ej., vasopresina). El rendimiento de aprendizaje empeora en ausencia de estas neurohormonas, o cuando su disponibilidad es reducida (cf. De Wied, 1973, 373 s.).
| Los factores como el elogio, la riña, el estrés de aprendizaje y la atención han demostrado su condición de refuerzos positivos o negativos; la cuestión es interesante para el proceso de aprendizaje motor y técnico. Su influjo se plasma en la mejora o la inhibición de la síntesis de proteínas. Así pues, el elogio y la riña pueden tener una expresión en formulas bioquímicas (cf. Kugler, 1981, 7). |
Las diferencias individuales en la capacidad de rendimiento memorístico, y por tanto de aprendizaje, se explican probablemente por la presencia variable de estas sustancias y por las consiguientes diferencias en los rendimientos de síntesis.
Como la fabricación sintética de estos “refuerzos de la memoria” es seguramente una cuestión de tiempo, suponemos que en un futuro próximo los rendimientos de aprendizaje motor serán muy elevados, mucho mejores desde el punto de vista del resultado, y ocuparán un tiempo mucho más reducido.
La teoría de los “bucles largos”
En el aprendizaje motor las células nerviosas del sistema nervioso central (neuronas) forman un reticulado específico a través de sus uniones sinápticas. El hecho tiene su expresión en la teoría de los “bucles del comportamiento” (cf. Hebb, 1949) o de los “bucles largos” (long loop; cf. Grimm/Nasher, 1978, 75 ss.) (fig. 3).
| Al inicio de un proceso de aprendizaje los aflujos de excitación (informaciones) tienen que recorrer varias veces el bucle como “circuito reverberador”, provocando los estímulos sucesivos necesarios para la memorización y fijando así el bucle (cf. Kugler, 1981, 5). |

basa en varios bucles, que se encadenan en diferentes planos anatómicos actuando de forma simultánea.
Dependiendo de la acción motora y del estado de rendimiento interaccionan diferentes bucles externos (p. ej., ópticos) e internos (p. ej., cinestésicos); un principiante controla sus movimientos más con la vista, y un “experto” más con las sensaciones cinestésicas (cf. Cratty, 1975, 412).
Basándonos en la “teoría de los bucles”, podemos definir de la siguiente manera el aprendizaje del movimiento y de la técnica:
| El aprendizaje induce la formación y
fijación de “bucles neuronales” específicos de los contenidos
informativos; a través de mecanismos propios de la memoria, estos bucles
se almacenan para un tiempo más o menos largo, y son por tanto
recuperables. La expresión “pulir un movimiento” adquiere desde este
punto de vista una dimensión fisiológica evidente. Desaprender (olvidar) significa la desaparición de un bucle motor anteriormente creado. El acto de reaprender se caracteriza por la sustitución de un bucle fijado por otro bucle más o menos parecido, pero en último término nuevo. |
Los bucles, según Grimm/Nasher (1978, 75), tienen un tiempo de existencia determinado por la fisiología. Se actualizan a través de estímulos externos específicos (entrada de información sensorial), de la experiencia o del control inconsciente. Una vez que se ha desarrollado un contenido determinado (output) desaparece el bucle, esto es, el circuito sistémico; las neuronas quedan libres para emprender otras tareas, esto es, quedan de nuevo disponibles para su integración en sistemas funcionales nuevos.
Sobre la base de las explicaciones previas se pueden explicar también, de manera parcial, las diferencias entre los individuos en cuanto a la velocidad del aprendizaje, y la razón por la que dicha velocidad disminuye con la edad. En el trasfondo causal se encuentra, desde un punto de vista neurofisiológico (esto es, dejando al margen los diferentes factores psicológicos, cognitivos y emocionales que influyen sobre el proceso del aprendizaje), la diferente capacidad de síntesis de hormonas y de proteínas.
Las diferencias en cuanto a la capacidad de aprendizaje se pueden apreciar también en las curvas de aprendizaje individuales.
Curvas de aprendizaje
Las curvas de aprendizaje son un método usual para visualizar la adquisición de habilidades motoras. Son representaciones gráficas del número de intentos y de los rendimientos conseguidos, y se consideran indicadores de la capacidad de aprendizaje y del transcurso de dicho proceso (cf. Cratty, 1975, 338/339; Singer, 1985, 36).
Dependiendo de factores como los métodos de entrenamiento, la distribución de las sesiones de entrenamiento, el tipo y nivel de la tarea o la edad del aprendiz, podemos obtener, como indica la figura 4, curvas diferentes para la misma tarea. Sobre el recorrido de las curvas influyen considerablemente una serie de factores favorables–entre otros, el tipo de enseñanza (demostración/imitación, explicación, permitir la experiencia o la sensación, v. también fig. 4), el comportamiento del pedagogo (paciente, benevolente, cooperativo, comprensivo, etc.), el entorno del aprendizaje (condiciones de aprendizaje y aparatos óptimos, entrenamiento con amigos, ejercicios de carácter lúdico, etc.)– o desfavorables para el rendimiento–pedagogo antipático (al que inconscientemente se considera incapaz), entorno de aprendizaje inadecuado (instalaciones deficientes, carencia de aparatos, condiciones climatológicas desfavorables, etc.) (cf. también fig. 5).
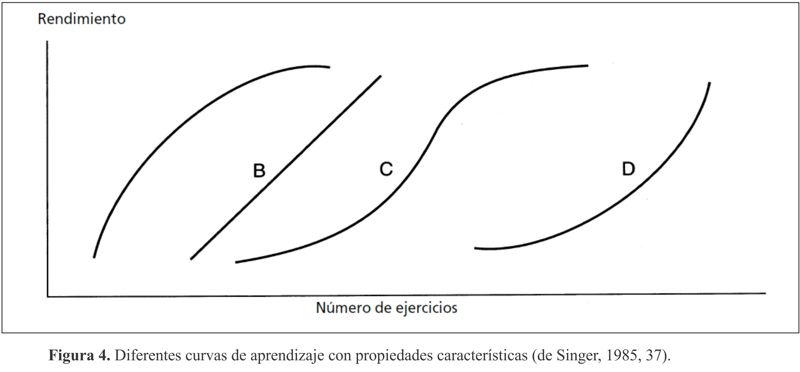
| Las curvas de aprendizaje “auténticas” son extremadamente difíciles de mantener. No existe “la” curva de aprendizaje. En el tipo de adquisición de la habilidad se refleja en todo momento la especificidad de la tarea o del aprendiz (cf. Singer, 1985, 36/37). |

En relación con el tipo de aprendiz, las curvas A y D podrían reflejar, respectivamente, un aprendizaje rápido y uno lento, mientras que en relación con la tarea podría tratarse de una tarea ligera y otra difícil/compleja; con esta última, el profesor debería tener la suficiente paciencia, y el alumno de aprendizaje rápido avanzaría rápidamente en la tarea fácil. En cualquier caso, el profesor tiene que garantizar una adaptación al proceso de aprendizaje individual.
Las estimaciones erróneas y los procedimientos equivocados condicionan el transcurso del aprendizaje o lo retrasan.
Artículos relacionados:
Fundamentos neurofisiológicos del aprendizaje de la técnica deportiva
Las Etapas en el Aprendizaje Motor, Neurociencias y DeporteI Simposio Internacional de Neurociencias aplicadas al Deporte

Bibliografía
Cratty, B. J.: Motorisches Lernen und Bewegungsverhalten. Limpert Verlag, Francfort, 1975.
Domagk, G. F., E. Schonne, G. Thines: “New experimental approaches to the inter-animal transfer of acquired information”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press, Nueva York – Londres, 1973, pp. 419-428.
Fjerdingstad, E. J.: “Chemical transfer of learned information in mammals and fish”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press, Nueva York – Londres, 1973, pp. 429-449.
Gay, R., A. Raphaelson: “Transfer of learning by injection of brain RNA: A replication”. Psychosomatic Science 8 (1967), 369-370.
Grimm, R. J., L. M. Nasher: “Long loop dyscontrol”. En: Desmedt, J. E. (ed.). Spinal and supraspinal mechanisrns of voluntary motor control and locomotion. Karger, Basilea. 1980.
Hebb, D. O.: The organization of behavior. Wiley & Sons, Nueva York, 1949.
Guttmann, H. N., G. Matwyshyn, M. Weiler: “Studies with dark avoidance and scotophobin”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press, Nueva York – Londres, 1973, pp. 391–317.
Huston, J. P., C. Mueller: “Memory facilitation by post-trial hypothalamic stimulation and other reinforces: a central theory of reinforcement”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS, Akademie Verlag, Berlín, 1979, pp. 175-186.
Kokonen, T.: “Implementation of associative memory in adaptive neural network”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag, Berlín 1979, pp. 43-53.
Kugler, J.: “Gedächtnis und Gedächtnisleistung neurophysiologisch beurteilt”. Sandorama (1981), 4, 5-9.
Lössner, B. y cols.: “The incorporation of leucine into protein of different rat brain structures during acquisition and consolidation of a ‘brightness discrimination’”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag, Berlín 1979, pp. 125-130.
Matthies, H.: “Biochemical regulation of synaptic connectivity”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press, Nueva York – Londres, 1973, pp. 531 ss.
Matthies, H., M. Krug, N. Popov (eds.): Biological aspects of learning, memory formation and ontogeny of the CNS. Akademie Verlag, Berlín, 1979.
Matthies, H.: “Biochemical, electrophysiological and morphological correlates of brightness discrimination in rats”. En: Brauer, M. A. B. (ed.). Brain mechanisms in memory and learning: from the single neuron to man. Raven Press, Nueva York 1979.
McConnell, J. V., D. H. Malin: “Recent experiments in memory transfer”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press, Nueva York – Londres, 1973, pp. 343-362.
Ott, T.: Mechanismen der Gedächtnisbildung. Fischer Verlag, Jena,1977.
Rahmann, H.: “The possible functional role for gangliosides in synaptic transmission and memory formation”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag, Berlín, 1979, pp. 83-110.
Singer, R. N.: Motorisches Lernen und menschliche Leistung. Limpert Verlag, Bad Homburg, 1985.
Sinz, R.: “Neurophysiologie und biochemische Grundlagen des Gedächtnisses”. En: Klix, F., H. Sydow (eds.). Zur Psychologie des Gedächtnisses. Huber Verlag, Berna, 1977.
Stark, H., T. Ott, H. Matthies: “Effects of neurohormones on memory consolidation”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag, Berlín, 1979, pp. 313-317.
Ungar, G.: “Evidence for molecular coding of neural information”. En: Zippel, H. P. (ed.). Memory and transfer of information. Plenum Press. Nueva York – Londres, 1973, pp. 317 ss.
Voronin, L. G., R. A. Danilova: “Protein metabolism and conformational changes in synaptic structures during learning”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag. Berlín, 1979, pp. 121-124.
Wied, D. de: “Peptides and behaviour”. En: Zippel, H. P. (ed.). Memory and transfer of information. pp. 373 385, Plenum Press, Nueva York – Londres 1973.
Wenzel, J., E. Kammerer, M. Frotscher: “Structural changes in the hippocampus after learning experiments”. En: Matthies, H., M. Krug, N. Popov (eds.). Biological aspects of learning, memory formation and ontogenity of the CNS. Akademie Verlag, Berlín 1979, pp. 361-364.