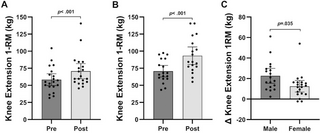
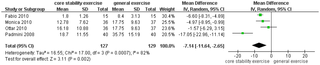
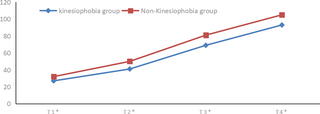
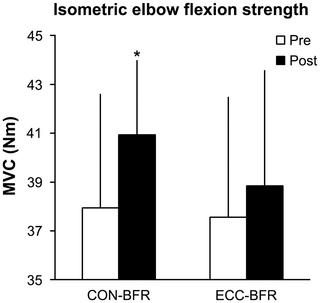
Resumen
Estudios recientes en pacientes que sufren de miopatías autoinmunes inflamatorias sugirieron que el entrenamiento de ejercicios moderado mejora o al menos estabiliza la fuerza y la función muscular sin inducir fallas de la enfermedad. Sin embargo, los mecanismos precisos involucrados en este efecto beneficioso no se han estudiado ampliamente. Aquí utilizamos un modelo de in vitro Mioblastos C2C12 estirados para investigar si el estiramiento mecánico podría influir en la proliferación de mioblastos o la expresión de genes proinflamatorios. Nuestros resultados demostraron que el estiramiento mecánico cíclico estimulaba el ciclo de células C2C12 y la regulación positiva temprana de las moléculas relacionadas con la vía del estroch mecánico en el músculo (calmodulina, NNOS, MMP-2, HGF y C-MET). Inesperadamente, el estiramiento mecánico también redujo la expresión de TLR3 y de proteínas que se sabe que representan autoantígenos en miopatías autoinmunes inflamatorias (MI-2, HRS, ADN-PKCS, U1-70). Curiosamente, la estimulación o la inhibición de las moléculas de calmodulina, NOS, HGF o C-Met in vitro afectó la expresión de autoantígenos y proteínas TLR3 confirmando su papel en la inhibición de autoantígenos y TLR3 durante el estiramiento mecánico. En general, este estudio demuestra por primera vez que el estiramiento mecánico podría ser beneficioso al reducir la expresión de autoantígenos musculares y de TLR3 proinflamatorio y puede proporcionar una nueva visión para comprender cómo el entrenamiento de resistencia puede reducir los síntomas asociados con la miositis.
Introducción
Los músculos esqueléticos responden bastante al estrés mecánico. La evidencia acumulada sugiere que varios componentes celulares que median la transducción mecánica están involucradas en la regulación de la activación de las células satelitales, iniciando la diferenciación miogénica y el desarrollo de los músculos esqueléticos (1–3). Se ha identificado el mecanismo molecular de activación y proliferación de células satelitales desencadenadas por estrés mecánico, y corresponde a una cascada de eventos iniciados por la formación del complejo de calmodulina de calcio (4), producción radical de óxido nítrico (no) (5,6), activación de metaloproteinasas de matriz (MMPS) (7), Liberación del factor de crecimiento de hepatocitos (HGF) con el segmento extracelular asociado de proteoglicanos, y la presentación posterior al receptor c-Met para generar una señal para la activación de las células satelitales (8–10).
En los pacientes que sufren de miopatías inflamatorias, el entrenamiento del ejercicio es, en la actualidad, no recomendarse, ya que se creía que la actividad física aumenta potencialmente la inflamación en los músculos afectados. Sin embargo, los estudios recientes en pacientes que padecen polimiositis (PM) y dermatomiositis (DM) respaldan la noción de que el entrenamiento del ejercicio moderado (p. Ej., 5 días a la semana para el programa de hogar de entrenamiento de resistencia de 12 semanas) mejora o al menos estabiliza la fuerza muscular y la capacidad funcional sin inducir enfermedades de la enfermedad ((((11). Se ha demostrado que el entrenamiento de resistencia restaura la función muscular al reducir la inflamación y la fibrosis tisular, mejorar la homeostasis metabólica a través de la expresión reducida de redes de genes proinflamatorios y profibróticos, y aumentando la expresión de genes de metabolismo oxidativo ((12). Posiblemente, el ejercicio moderado al inducir la tensión muscular podría inducir la regulación positiva de las moléculas clave involucradas en la respuesta al estrecho mecánico que culminó en la activación de las células satelitales y en la reparación muscular parcial. Sin embargo, las consecuencias de la regulación positiva de estas moléculas en la expresión de proteínas que se sabe que representan autoantígenos potenciales o en genes proinflammatoy aún no se han explorado. Como la regulación positiva concomitante de los receptores tipo toll (TLR) o de los autoantígenos potenciales podría agravar la enfermedad al estimular las citocinas y la producción de quimiocinas, así como la activación de las células T autoinmunes (13–17), pensamos aquí para explorar cómo la inducción de la vía del estiramiento mecánico podría influir en su expresión.
Esto es de particular importancia en situaciones inflamatorias crónicas, ya que las células musculares regenerativas se consideran la principal fuente de autoantígenos y expresan niveles más altos de TLR en comparación con los miotubos diferenciados maduros. De acuerdo con esto, mioblastos cultivados in vitro expresar altos niveles de autoantígenos y de TLR3 y TLR7 que son notablemente regulados a medida que las células se diferencian en miotubos (15,16). Exploramos aquí las consecuencias del estrés mecánico con respecto a la proliferación de mioblastos y la expresión de autoantígenos y de TLR3 y TLR7 en un in vitro modelo de estiramiento mecánico. Para eso, los mioblastos C2C12 se cultivaron y se sometieron a estiramiento mecánico controlable durante el período cíclico utilizando el sistema FlexCell. Curiosamente, nuestros resultados muestran que el estroch mecánico estimuló, como se esperaba, el ciclo celular, pero notablemente también redujo la expresión de TLR3 y de proteínas que se sabe que representan autoantígenos potenciales. Por lo tanto, estos datos pueden ayudar a comprender mejor el papel beneficioso recién reconocido del ejercicio moderado en pacientes que padecen miopatías inflamatorias.
Materiales y métodos
Cultivo celular e inducción de estrach mecánico
Las células C2C12 (ATCC, EE. UU.) Se colocaron en placas en la mezcla de nutrientes mediana del águila modificada de Dulbecco, la mezcla de nutrientes medias F-12 (DMEM/F12, Thermo), suplementado con suero bovino fetal al 10%, FBS), 100Units/ml de penicilina y 100 mg/ml de sulfato de estrepticina en A 5% Co Co Co Co CO a 5%2-Cámara humidificada (Heraeo, Alemania) a 37 ° C. Las células se cultivaron a aproximadamente 70-80% de confluencia y se usaron para los próximos experimentos.
Para la inducción del estiramiento mecánico, las células C2C12 se resuspendieron en el medio de crecimiento (GM) definido anteriormente y se colocaron en placas en las seis placas de pozos de fondo flexible recubierto de colágeno tipo I (colágeno de placas Bioflex I, Flexcell International Corporation, Hillsborough, NC, EE. UU.) Y se incubaron a 37 ° C en una co Co Co Co2 Incubadora durante 24 h antes de aplicar la tensión mecánica. El número de células sembradas se limitó a 1 × 105 células/bueno, para mantener la confluencia celular en menos del 45% durante las primeras 24 h de cultivo. Luego, las células se sometieron a una tensión cíclica de un alargamiento del 10%, a una frecuencia de 0.25Hz durante 2 h por día, o de alargamiento del 15%, con frecuencia de 1.0Hz, durante 1H por día, utilizando un aparato de estiramiento de vacío controlado por computadora (Sistema de tensión FX-5000, FlexCell International Corporation). En los cultivos de control, las células fueron un crecimiento en paralelo con condiciones experimentales idénticas, pero se dejaron sin estirarse. Las células C2C12 se evaluaron 2, 4 o 6d después del inicio de la cepa cíclica para evaluar su tasa de ciclo o la expresión de moléculas en el estudio. Como notamos signos de diferenciación a los miotubos después del día 4, y para evitar cualquier efecto de confusión debido a la diferenciación celular, los mioblastos siempre se estiraron por menos de 4 días.
Análisis del ciclo celular utilizando citometría de flujo
Para el análisis del ciclo celular, las células C2C12 estiradas y no estiradas se recogieron y se fijaron con etanol al 80% a 4 ° C durante 24 h, y se incubaron aún más durante la noche a 4 ° C con 1 ml de una mezcla de tinción de propidio (PI) de acuerdo con el protocolo del fabricante (KIT de GenMed Scientifics Inc.). Después de manchar, 105 Las células se analizaron por citometría de flujo utilizando un citómetro de flujo de FACS Calibur (BD, EE. UU.). El índice de proliferación de ADN relativo (DPI = (s%+g2/M%)/(s%+g2/M%+g0/GRAMO1%)) se usó para evaluar la tasa de ciclo de las células C2C12.
Uso de agonista o antagonista para interferir con las moléculas clave involucradas en la respuesta al estiramiento mecánico
En algunos experimentos, las células fueron tratadas con agonistas o antagonistas para interferir con las moléculas clave involucradas en la respuesta al estiramiento mecánico. Para eso, los cultivos de células C2C12 se lavaron a 24 h después del plato con DMEM sin suero y luego se trataron durante 4 h con las siguientes moléculas agonistas o antagonistas: ionóforo de calcio A23187 (3 µM, Calbiochem-Novabiochem (La Jolla, CA)); etilenglicol tetra-acetato (egta, 1.8 mm, tocris, británico); cloruro de calmidazolio (R24571, 0.3um, Santa Cruz, EE. UU.); HGF murino recombinante (10ng/ml, I + D); Conejo anti-HGF policlonal (2UG/ml, abcam); HGF R (c-Met, 1UG/ml recombinante; Conejo anti-C-C-Met (2UG/ml, Santa Cruz, EE. UU.); MMP-2 humano recombinante (cf, 10ng/ml, R&D, EE. UU.); Inhibidor de MMP-2 I (250ng/ml, Santa Cruz, EE. UU.); deshidrato de nitroprusiato de sodio (SNP, 8ug/ml, sigma); o L-NG-Nitroarginina Metil Ester (L-Name, 10um, Santa Cruz, EE. UU.). Luego se lavaron las células con DMEM sin suero y se cosecharon para su análisis por QRT-PCR o Western Blot. Las células C2C12 cultivadas en DMEM libre de suero durante 4H pero sin la adición de agonista o antagonista se tomaron como controles.
Aislamiento y análisis de ARN por RT-PCR cuantitativa en tiempo real
24 horas después del cese del estiramiento, o 2 horas después del tratamiento con agonistas o antagonistas, se extrajo ARN total de las células C2C12. Para eso, las células se cosecharon en 1 ml de TRIZOL® (Invitrogen) y el ARN total se purificaron de acuerdo con las instrucciones del fabricante. Luego se usó 1 μg de ARN total para la transcripción inversa (RT) con el kit comercialmente disponible (retaid de primer kit de síntesis de ADNc de cadena, fermentas). La reacción en cadena de la polimerasa en tiempo real (PCR) se realizó por triplicado con un sistema ABI Stepone Plus (Applied Biosystems, EE. UU.) Y un kit de mezcla SYBR Green/Rox QPCR de SYBR marcado con fluorescencia (fermentosas) para los siguientes genes: MI-2, HARS, ADN-PKCS, U1-70, TLR3 y TLR7, y con con con con respecto gliceraldehído-3-fosfato deshidrogenasa (GAPDH) tomada como un control endógeno (secuencias de cebadores y tamaños de amplicones se enumeran en Tabla 1). Los resultados se analizaron con el software SOS2.1 (Applied Biosystems). La expresión de los genes se calculó a partir del ciclo de umbral preciso (TC), que es el ciclo de PCR al que primero se puede detectar un aumento en la fluorescencia de las sondas verdes SYBR por encima de la señal de referencia. Los valores de CT para GAPDH se compararon con los de MI-2, HARS, ADN-PKCS, U1-70, TLR3 y TLR7, y en cada pozo para calcular ΔCT. Los datos de las condiciones tratadas se expresaron en relación con la señal obtenida para el promedio de los controles no tratados por el cálculo de ΔΔCT. Se promediaron los valores de ΔΔCT triplicado para cada muestra.
| Genes | Secuencia (5 ‘→ 3’) | Tamaño de amplicón |
|---|---|---|
| Mi-2 | Para Cccgaggagtgtgga acta | 62 pbp |
| Rdo Ccctaccaccctagccaag | ||
| Hars | Para Gatgggatgtttgctgtctg | 114 pbp |
| Rdo TCCCACCATCTCATTCTTCA | ||
| ADN-PKCS | Para Atatccttggcaggacttgg | 92bp |
| Rdo Aggtcctccggagacagaa | ||
| U1-70 | Para Gacagcagtaagtggagaca | 91bp |
| Rdo Gcacgaacagagtcttgaa | ||
| TLR3 | Para Tctgttgcgaagaggaatg | 114 pbp |
| Rdo Aattccgagatccaagtgct | ||
| TLR7 | Para Ttgcaactgtgatgctgtgt | 106bp |
| Rdo Tttgtgtgctcctggaccta | ||
| Gapdh | Para Caatgtgtcgtcgtggatct | 124 pbp |
| Rdo Gtcctcagtgtagcccaagatg |
Tabla 1. Secuencias de cebador para QRT-PCR.
Análisis de transferencia Western
Las células se recogieron, se lavaron con PBS helado y la extracción de proteínas se realizó de acuerdo con el protocolo del fabricante (Keygen; China). Las concentraciones de proteínas se evaluaron utilizando un kit de ensayo BCA (Solarbio, Beijing, China). Cantidades iguales de proteínas se electroforesis en gel de 6-12% SDS-poliacrilamida y se transfirieron a la membrana PMobilon P (Millipore, EE. UU.). Las membranas se bloquearon en leche seca al 5% no grasas en solución salina tamponada con Tris/Tween-20 (TBS-T: Tris de 20 mm, PH7.5, NaCl 150 mM, 0.05% Tween-20) durante 1 h a temperatura ambiente. Se usaron el siguiente anticuerpo policlonal de conejo o cabra para la detección: anti-MI-2 (1: 2000, ABCAM); anti-hars (1: 400, abcam); Anti-DNA-PKCS (1: 100, Santa Cruz); anti-U1-70 (1: 100, Santa Cruz); anti-TLR3 (1: 800, abcam); anti-TLR7 (1: 100, Santa Cruz); antialmodulina (1: 100, Santa Cruz); anti-nna (1: 100, Santa Cruz); anti-HGF (1: 400, abcam); anti-C-Met (1: 100, Santa Cruz); anti-MMP-2 (1: 400, abcam); Anti-GAPDH (1: 3000, Kangchen, China). Los anticuerpos policlonales primarios se incubaron durante 20 h a 4 ° en leche seca sin grasa al 5% en TBS-T. La membrana se lavó luego tres veces en TBS-T y se incubó durante 1 h a temperatura ambiente con un anticuerpo de cabra conjugada de cabra o burro de cabra o burro de 1: 4000, con la leche seca sin grasa 5% en Tbs-T. Después de tres lavados en TBS-T, las bandas de proteínas se visualizaron por quimioluminiscencia mejorada …