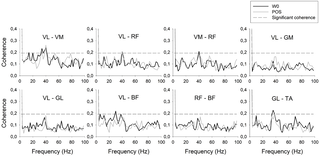
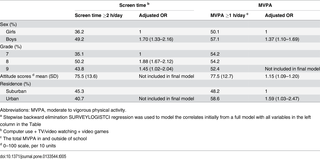
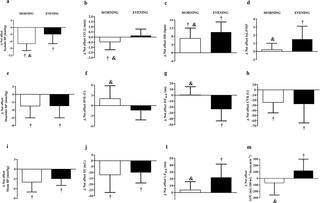
Resumen
Los canales de calcio dependientes de voltaje de tipo P/Q (VDCC) son esenciales para la transmisión sináptica en las uniones neuromusculares de mamíferos adultos (NMJ); Sin embargo, la ubicación subsináptica de los VDCC en relación con las zonas activas en NMJ de roedores, y la modificación funcional de VDCC por la interacción con el fagot de la proteína de zona activa sigue siendo desconocida. Aquí, mostramos que los VDCC P/Q-Type se distribuyen en un patrón punteado dentro de los terminales presinápticos NMJ y se alinean en tres dimensiones con fagot. Este patrón de distribución de VDCC de tipo P/Q y fagot en NMJS es consistente con nuestro estudio anterior que demuestra la unión de VDCC y fagot. Además, ahora mostramos que la interacción entre los VDCC de tipo P/Q y el fagot suprimió significativamente la propiedad de inactivación de los VDCC de tipo P/Q, lo que sugiere que la CA2+ La afluencia puede aumentar el fagot para una transmisión sináptica eficiente en NMJS. Sin embargo, el nivel de fagot presináptico se atenuó significativamente en NMJ de ratas envejecidas, lo que sugiere una atenuación de la función VDCC debido a la falta de esta interacción entre VDCC y fagot. Es importante destacar que el nivel de fagot disminuido en NMJ envejecido se vio mejorado por el entrenamiento de fuerza isométrica de los músculos durante dos meses. El entrenamiento aumentó la inmunorreactividad del fagot en NMJ sin afectar el tamaño de la sinapsis. Estos resultados demostraron que los VDCC de tipo P/Q se acumulan preferentemente en las zonas activas de NMJ y juegan un papel esencial en la transmisión sináptica junto con el fagot de la proteína de la zona activa. Este mecanismo molecular se ve afectado por el envejecimiento, lo que sugiere una función sináptica alterada en NMJ envejecidos. Sin embargo, el nivel de fagot en NMJ envejecido puede mejorarse mediante el ejercicio muscular.
Introducción
Transmisión sináptica en los Iniciados de NMJS para adultos por la CA2+ afluencia a través de los VDCC de tipo P/Q (1), (2) y fusión de vesículas sinápticas en las zonas activas (3). Según los estudios de NMJ y otras sinapsis, se estima que los VDCC esenciales para la transmisión sináptica se localizan en o en las cercanías estrechas de las zonas activas (2), (4)–(8). Una confirmación anatómica de estos análisis es más adecuada en grandes sinapsis como las NMJ de mamíferos, pero no se ha revelado la ubicación relativa de los VDCC de tipo P/Q y las zonas activas de NMJ. Los estudios de inmunohistoquímica publicados por otros muestran una distribución relativamente difusa de VDCC de tipo P/Q que cubren todos los terminales presinápticos de NMJ de roedores (9)–(12). Esta distribución difusa de VDCC de tipo P/Q en los terminales presinápticos es algo inesperada teniendo en cuenta la distribución discreta y punzante de zonas activas en NMJ de roedores detectados por microscopía electrónica (13) e inmunohistoquímica (14)–(16). Por lo tanto, primero preguntamos si los VDCC P/Q-Type se localizan en las zonas activas de NMJ. En relación con la acumulación de VDCC en zonas activas, nosotros y otros hemos demostrado que los VDCC y las proteínas de la zona activa forman complejos de proteínas (14), (17)–(24). Hemos demostrado que la subunidad VDCC ß y el fagot interactúan para organizar las zonas activas de NMJ (14). Sin embargo, no se conoce el efecto de la interacción entre el VDCC de tipo P/Q en la función del canal. Por lo tanto, probamos los VDCC P/Q-Type utilizando la grabación de pinzas de parche y demostramos que la interacción de VDCC y fagot de Type P/Q modifica la función VDCC. Esta modificación tiene el potencial de desempeñar un papel importante en la transmisión sináptica en NMJS. Si esta interacción es esencial para la transmisión sináptica de NMJ, nuestros hallazgos recientemente de niveles de proteína de fagot atenuado en NMJ de ratones envejecidos pueden tener efectos perjudiciales en la función NMJ (15). Esta visión es consistente con las alteraciones fisiológicas registradas en NMJS envejecidos por otros (25)–(27) y puede estar relacionado con la denervación de NMJ envejecidos (25), (28)–(30). Por lo tanto, nos llevó a buscar formas de mejorar la pérdida de fagot en los NMJ envejecidos. Intentamos hacer ejercicio roedores envejecidos porque los efectos beneficiosos de la intervención del ejercicio para el sistema nervioso se han descrito anteriormente (31)–(35). Identificamos que el nivel de fagot se puede recuperar en NMJ envejecido por el entrenamiento muscular.
Resultados
P/Q-Type VDCCS localizarse en las zonas activas de NMJ
El punto de partida para este estudio fue nuestro hallazgo previo de que los VDCC presinápticos son esenciales para organizar zonas activas, y funcionan como proteínas de andamiaje que anclan las proteínas de zona activa en los terminales presinápticos. (14), (16). En estos estudios, hemos demostrado que los VDCC utilizan su dominio citosólico para unirse a las proteínas de zona activa, que se localizan como punciones pequeñas discretas en NMJ. Sin embargo, la ubicación relativa de los VDCC de tipo P/Q y las proteínas de la zona activa no se han analizado en los estudios de inmunohistoquímica publicados por otros. (9)–(12). Además, estos patrones de tinción de VDCC de tipo P/Q en NMJ fueron diferentes del patrón de tinción punteada discreta de las proteínas de zona activa que identificamos (14)–(16). Por lo tanto, comenzamos examinando el patrón de distribución de VDCC de tipo P/Q en NMJS. Nos centramos en los VDCC de Type P/Q porque los NMJ para adultos utilizan principalmente este VDCC para la transmisión sináptica (1).
Un anticuerpo comercial contra P/Q-Q-Type VDCCS nmjs de ratones de tipo salvaje en el día 15 postnatal en un patrón punteado (Fig. 1A). Es importante destacar que estas señales estaban ausentes en los nmjs de ratones Knockout VDCC de littermate P/Q-Q-Type (CACNA1A-/ –)demostrando la especificidad de las señales de inmunohistoquímica (Fig. 1A). La reconstrucción tridimensional utilizando las pilas Z del microscopio confocal reveló que los VDCC de tipo P/Q se distribuyen en un patrón punteado discreto dentro de los NMJ marcados por α-bungarotoxina que se une específicamente a los receptores de acetilcolina (Fig. 1B, C). En la vista de la sección transversal ortogonal del NMJ, se detectaron VDCC de tipo P/Q en la parte inferior de la canaleta primaria de las placas finales, donde residen los terminales del nervio motor (Fig. 1B).

(A) VDCC de tipo P/Q teñidos con un anticuerpo contra la subunidad α (Cav2.1, verde) en NMJ del músculo esternomastoideo de los ratones de tipo salvaje del día 15 postnatal teñidos con Alexa Fluor 594-bungarotoxina marcada con etiqueta para etiquetar los receptores acetilocolinos (ASHR, rojo). La inmunorreactividad está ausente en el área NMJ de los ratones Knockout VDCC de type P/Q-Q-type ((CACNA1A-/ –) demostrando la especificidad de las señales inmunohistoquímicas. El mismo resultado se confirmó en cuatro camadas independientes. (B) Se detectaron VDCC de tipo P/Q dentro del canalón primario de las placas finales marcadas con α-bungarotoxina, donde residen los terminales del nervio motor (puntas de flecha). Se muestra una vista YZ-ortogonal (un solo plano óptico) de una región indicada por una línea naranja en A. El nervio se coloca hacia la parte superior del eje Z y el músculo hacia la parte inferior. También se detectaron algunas señales VDCC de tipo P/Q en el lado muscular. (C) VDCC de tipo P/Q alineados con el fagot de la proteína de zona activa en NMJ (puntas de flecha). El músculo esternomastoideo del día 21 postnatal 21 ratones de tipo salvaje se tiñó usando un anticuerpo de subunidad α de tipo VCCC anti-P/Q (rojo), un anticuerpo anti-Bassoon (verde) y la α-bungarotoxina marcada con Alexa Fluor 647 (azul). La visión XY máxima proyectada de la pila Z confocal se muestra en C1. Paneles c2 y C3 Mostrar vistas ortogonales YZ y XZ (un solo plano óptico) en las posiciones indicadas por las líneas naranjas en C1. Paneles c4 y C5 mostrar una región magnificada de c1 indicado por la caja de puntos (c4) y línea naranja dentro de la caja punteada (c5). Muchas señales VDCCS de fagot y P/Q type (puntas de flecha blanca) se alinean en las vistas XY (C4) e vistas ortogonales yz (c5plano óptico único). Cª5el nervio se coloca hacia la parte superior, y el músculo se coloca hacia la parte inferior. (DO6) Análisis de colocalización del fagot y P/Q-Type VDCC dentro de los terminales presinápticos NMJ de ratones de tipo salvaje del Día Postnatal 21 por los coeficientes de los Manders (M). Cierto grado de colocalización de estas proteínas se indicó por los valores de los coeficientes de los manders significativamente más altos para la superposición de fagot con P/Q-Type VDCC (0.42 ± 0.04, 3 nmjs, BSN en PQ) y P/Q-Type VDCC Overplaon con axeto (0.39 ± 0.02, 3 NMJS, PQ, Bause) para Bailon) a comparación con Bailon) a comparación con Bailon) a comparación con BaLoon). receptor de acetilcolina superpuesto mínimamente con neurofilamento (0.11 ± 0.03; 6 nmjs, AChR en NF). Se detectó una diferencia significativa utilizando ANOVA unidireccional (P = 0,0002). Los asteriscos indican diferencias significativas contra AChR en NF por Bonferroni Post-Test. Barras de escala: A, 10 µm; B, C, 1 µm.
Es importante destacar que muchas señales punteadas de VDCC de tipo P/Q que se superponían con la tinción de α-bungarotoxina alineada con el patrón de distribución punteada del aicio de la proteína de la zona activa como se muestra en las vistas de arriba hacia abajo y las vistas ortogonales de las pilas z confocales (((Fig. 1C1–5). El patrón de tinción punteada de VDCC y fagot de tipo P/Q fue diferente del patrón de tinción inmunohistoquímico difuso de la proteína SV2 asociada a la vesícula sináptica SV2 (14)–(16)sugiriendo su localización en las zonas activas. Este patrón de distribución de P/Q-Type VDCC y fagot en NMJS es consistente con nuestro estudio anterior que demuestra la unión de VDCC y fagot (14). De la puncta VDCC P/Q-Q-Q-Type identificada manualmente que se superpuso con la señal de α-bungarotoxina, el 85% de la puncta tenía una señal de fagot superpuesta que sugiere que el fagot es unión al lado citosólico de P/Q-Type VDCC en terminales presinápticos ((Fig. 1C4). Furthermore, colocalization of P/Q-type VDCC and Bassoon was indicated by the significantly higher Manders’ coefficient M values for Bassoon overlapping with P/Q-type VDCC (0.42±0.04) and P/Q-type VDCC overlapping with Bassoon (0.39±0.02) compared to the M value for acetylcholine receptor overlapping minimally with neurofilamento (0.11 ± 0.03) (Fig. 1C6). Estos datos demostraron que los VDCC de tipo P/Q se localizan en los terminales presinápticos de NMJ con el fagot de la proteína de zona activa.
Curiosamente, se observaron algunas señales VDCC de tipo P/Q cerca del grupo de receptores de acetilcolina (Fig. 1). Esta observación es compatible con la inmunorreactividad VDCC de tipo P/Q de Schwann en células Schwann (10) y músculos (36).
La interacción con fagot modula la inactivación de VDCCS
P/Q-Type VDCCS y fagoton preferentemente co-localizados en los NMJ, y previamente demostramos que estas dos proteínas se unen directamente (14). Por lo tanto, un papel potencial de esta interacción en el terminal presináptico de NMJS es modificar las funciones VDCC para mejorar la transmisión sináptica. Para probar esta hipótesis, analizamos las características del canal de calcio con o sin fagot. Una línea celular que expresa establemente el VDCC de tipo P/Q (Cav2.1, ß1a, α2/δ) se transfectó con un vector bicistrónico que expresa fagot y GFP. Las células transfectadas se identificaron por la fluorescencia GFP y se analizaron mediante la técnica de pinza de parche de células enteras. Las células transfectadas con un vector bicistrónico que expresa solo GFP se usaron como controles.
El efecto más destacado del fagot en los VDCC se observó en los parámetros de inactivación. La propiedad de inactivación de P/Q-Type VDCC fue suprimida significativamente por el fagot coexpresado (Fig. 2a, Tabla 1). El potencial de medias inactivación se cambió por 6.1 mV en células que expresan fagot (−40.0 ± 1.7 mV) en comparación con los controles (−46.1 ± 0.6 mV). Observamos un cambio de despolarización significativo en la dependencia del voltaje de la inactivación. Este efecto de fagot en P/Q-Type VDCC es similar al efecto de RIM1 en VDCCS (17), (19).
(A) Izquierda, inactivación de las corrientes VDCC (Cav2.1) de Type P/Q en células BHK que expresan de manera estable VDCC y se transfectan con un vector de expresión PBassoon-IRES2-GFP (BSN, rojo) o un vector Pires2-GFP vacío (control, negro). Las amplitudes máximas se normalizaron para BA2+ Corrientes provocadas por pulsos de 2 s a 0 mV desde un potencial de retención de –100 MV. Gráfico correcto …