
1Club del Personal de Banco Mendoza.
2Asociación Amateurs de Hockey Sobre Césped de Mendoza.
Resumen
INTRODUCCIÓN
Es ampliamente discutida la temática referida a cómo se deben generar adaptaciones aeróbicas que desarrollen la capacidad de resistencia física. La discusión se agudiza en el marco de los deportes de conjunto de dinámica intermitente donde es conocida la relevancia del estímulo de alta intensidad (por encima de la VAM (Velocidad Aeróbica Máximao velocidad del VO2máx) hasta «all out» o máxima intensidad posible) para la producción de adaptaciones sobre los sistemas anaeróbicos de producción energética y las fibras rápidas (tipo II) (Cometti, 2002). Parece reconocerse este hecho pero como en este tipo de disciplinas el metabolismo aeróbico también es determinante, en especial por su papel en la recuperación entre las distintas acciones y carrera (Barbero Álvarez et al., 2006a y b), surge la discusión sobre cómo estimular la capacidad de resistencia teniendo en cuenta todos estos aspectos. Durante décadas se ha establecido una directa relación entre los trabajos de intensidades bajas y moderadas con el estímulo adaptativo de la capacidad de resistencia, casi desechando la posibilidad de que las metodologías de altas intensidades también sean pertinentes para el estímulo de dicha capacidad. Es por esta razón que es conocido como a la hora de periodizar el entrenamiento de la capacidad de resistencia se sostiene en general que lo ideal en este aspecto sería realizar un primer periodo de base donde se priorizarían metodologías de intensidades inferiores a la VAM para luego de este período avanzar sobre el estímulo más intenso y específico (Navarro Valdivieso et al., 1999). Por supuesto que estas metodologías de “base”, serían también de una menor especificidad si ingresamos en el contexto de los deportes de conjunto de dinámica intermitente. Este paradigma señalado junto con el modelo anteriormente descripto sobre la necesidad de metodologías de base de intensidades bajas y más inespecíficas previas a otros trabajos ha sido desestimado por la literatura actual abriendo nuevos tópicos de discusión y paradigmas en relación a la temática en cuestión (Astrand et al., 1960a; Barbero Álvarez et. al., 2005; Cometti, 2002; Morera Siercovich, 2005; Kohan, 2008).
Esto se debe a que distintas investigaciones han ido demostrando la relación directa que existiría entre la alta intensidad y la resistencia y las adaptaciones aeróbicas, entre los sistemas bioenergéticos anaeróbicos y el aeróbico. Esta relación es evidenciada a través de estudios descriptivos-correlacionales, que relacionan la alta intensidad, medida a través de los metros a alta intensidad recorridos durante la competencia, con la resistencia aeróbica, medida a partir de distintos protocolos evaluativos para la resistencia como el YOYO de recuperación intermitente (utilizando como indicador los metros recorridos durante dicho protocolo evaluativo), con un r = 0.75 (P < 0.05) (Krustrup & Bangsbo, 2001) y r = 0.71 (P < 0.05) (Krustrup et al., 2003). Una revisión también señala un r = 0.70 (P < 0.05) entre estas variables (Bangsbo et. al. 2008). En esta línea también se han encontrado significativas correlaciones entre sprints repetidos y utilizados indicadores de la potencia aeróbica como lo es el VO2máx, como la encontrada entre el tiempo medio de los sprints en un test de RSA y el VO2máx (P < 0.05) (da Silva et al., 2010). Entonces en este sentido, un análisis a realizar a partir de este tipo de estudios es que si la alta intensidad, medida en el campo de juego a través de los metros a alta intensidad recorridos (>15 km.h-1) o en un protocolo de sprints repetidos, correlaciona significativamente y/o en una proporción elevada con evaluaciones de la resistencia aeróbica altamente utilizadas en el marco de los deportes de conjunto (YOYO de recuperación intermitente) y reconocidos parámetros aeróbicos como el VO2máx, indicaría entonces que entre la alta intensidad y la resistencia aeróbica existe una directa relación, quedando en última instancia para la discusión aún abierta el terminar de establecerse la proporción de la misma.Por lo tanto se puede deducir entonces que utilizando protocolos de entrenamiento de altas y máximas intensidades también se podrían inducir adaptaciones aeróbicas, algo que se observa ya directamente en diversos estudios que a continuación se señalarán.
Diversos estudios encontraron que los entrenamientos de altas intensidades produjeron significativos incrementos en los resultados de distintos tipos de protocolos evaluativos de la resistencia (Stepto et al., 1999; Burgomaster et al., 2005; Krustrup & Bangsbo, 2001; Ferrari Bravo et al., 2008). Incluso luego de estos procesos de entrenamientos intensos se evidencian significativos incrementos en la capacidad de resistencia medida en forma directa por los metros a alta intensidad (>15 km·h-1)recorridos durante la competencia (Krustrup & Bangsbo, 2001).
Siguiendo esta línea, son variados también los estudios que a partir de distintos parámetros de la resistencia física demuestran las adaptaciones aeróbicas que generan distintos protocolos de entrenamientos fraccionados deintensidades máximas, evidenciadas en elevaciones en el VO2máx (Mac Dougall et al., 1998; Rodas et al., 2000; Ferrari Bravo et al., 2008), en el aumento de la densidad y actividad de enzimas aeróbicas (Mac Dougall et al., 1998; Rodas et al., 2000; Burgomaster et al., 2005; Burgomaster et al., 2007; Gibala et al., 2006; Burgomaster et al., 2008), en el incremento de los capilares por fibra (capilarización fibrilar) (Jensen, L. et al., 2004) y en el proceso de biogénesis mitocondrial que se estimularía luego de dichos procesos (Little et al., 2010).
Es por este marco científico que enespecial en el contexto de los deportes de conjunto de dinámica intermitente (fútbol, rugby, hockey s/césped, etc.) la investigación está dando bases para el replanteo sobre ese modelo popularizado que plantea la necesidad de utilizar períodos de tiempo en entrenamientos más inespecíficos de intensidades bajas y altos volúmenes previos a otros tipos de protocolos más intensos y específicos, con la fundamentación de que estos generarían las llamadas “bases aeróbicas”, ya que como los estudios demuestran dichas «bases aeróbicas» pueden irse formando también, y quizás hasta en forma más rápida, eficaz y completa, directamente con diversos protocolos de entrenamientos más intensos y/o específicos. La presente revisión persigue como objetivo principal presentar y analizar información científica que sirva de sustento para este replanteo y que contribuya a continuar con esta discusión.
CARACTERÍSTICAS DE LAS METODOLOGÍAS FRACCIONADAS: INTERVALADOS, INTERMITENTES Y SPRINTS REPETIDOS O “REPEATED SPRINT ABILITY” (RSA)
En los escritos de habla sajona en torno a la concepción del entrenamiento intervalado y el intermitente, los diversos autores utilizan diferentes terminologías para referirse a los mismos, incluso las diferencias se observan cuando comparamos estos escritos con la mayoría de habla hispana. Es por esta razón que resulta difícil realizar definiciones y diferenciaciones de estas metodologías. A modo de resumen, y en busca de un estandarización general, se tomará como base el análisis sobre la temática que realiza Casas, A. (2008) donde basado en un importante sustento bibliográfico, con autores que han sido pioneros en este abordaje (Astrand, I., Astrand, P.O., Daniels, J., Scardina, N., Nöcker, J., Bohlau, N., Hohnen, R.), expone que el entrenamiento intermitente (“Intermittent Training”) básicamente consta demomentos de trabajo/pausa más cortos que el entrenamiento intervalado (“interval training”). En el caso del intermitente los tiempos de trabajos y de pausas no superarían el minuto, incluso se podría hablar de tiempos no superiores a los 15 o 20 s (5 s x 5 s, 10 s x 10 s, 15 s x 15 s, 20 s x 20 s, 10 s x 20 s, etc.).
Por otro lado el trabajo intervalado conocido como “método clásico de Friburgo” posee características bastante diferentes, con tiempos de trabajo/pausa más largos (de 1’ a 5’), es un entrenamiento tipo “por pasadas”(pasadas de 200, 300, 400, 600, 800, 1000 metros), con relaciones trabajo/pausa más largas (1, 15 min x 1, 15 min; 2 x 2 min; 3 x 3 min; 1, 30 min x 3 min; etc.) (Casas A., 2008).
En cuanto a la intensidad, si se toma como parámetro los porcentajes en relación a la vVO2máx o la VAM, en el intervalado la intensidad variaría entre el punto de crisis metabólico de repercusión sistémica (conocido tradicionalmente como “umbral anaeróbico”) y la VAM (ejercicios submáximos a máximos) y en el intermitente partiría del 100% de la VAM en adelante (ejercicios máximos a supramáximos si tomamos a la VAM como parámetro de referencia) (Casas, A., 2008).
Por último la metodología de sprints repetidos relacionadas con la capacidad de repetir sprints o RSA (Repeated Sprint Ability) es semejante a la metodología intermitente con la diferencia de que en la metodología de sprints sucesivos la intensidad es siempre la máxima posible para cada esfuerzo, por lo que permitiría menores volúmenes totales. Entonces, la diferencia principal radica en la intensidad, donde para las metodologías relacionadas con la RSA será siempre máxima (“all out”), la cual por lo general se encuentra por encima del 150% de la VAM aproximadamente (Casas A., 2008). Es por esto que si tomamos como parámetro la velocidad máxima de los sujetos, entonces al intermitente lo podríamos catalogar como una metodología de intensidades submáximas a diferencia de los RSA que con este criterio se catalogaría como una metodología de intensidades máximas.
En la bibliografía científica encontramos básicamente 2 tipos de protocolos de entrenamientos de sprints repetidos, uno denominado por la bibliografía como sprints intermitentes o sprints intervalados (Sprint interval training) y otro denominado como sprints repetidos propiamente dicho (RSA). El primer protocolo nombrado consta básicamente de esfuerzos un poco más extensos con pausas completas (20 s a 30 s de esfuerzo x 2, 30 a 4 min de pausa) (Mac Dougall et al., 1998; Burgomaster et al., 2005; Burgomaster et al., 2007; Gibala et al., 2006; Burgomaster et al., 2008) y la segunda consta de esfuerzos más cortos de menos de 15 s, generalmente entre 5 y 10, con pausas semi-completas por serie que rondarían entre 3 y 6 veces el tiempo de trabajo (densidad 1/6 a 1/3) (Rodas et al., 2000; Impellizzeri et al., 2006; Ferrari Bravo et al., 2008; Aziz et al., 2008; Impellizzeri et al., 2008; da Silva et al., 2010).
ADAPTACIONES AERÓBICAS GENERADAS POR ESFUERZOS REPETIDOS DE INTENSIDADES MÁXIMAS (INTERMITENTES DE MÁXIMA INTENSIDAD, INTERVÁLICOS DE SPRINT, SPRINTS REPETIDOS O RSA)
En la tabla tomada y adaptada de Mac Dougall et al. (2004) (Tabla 1) se detalla a modo de resumen estudios que evidencian la relación entre los esfuerzos repetidos de intensidades máximas, de sprints repetidos (RSA), y las adaptaciones aeróbicas. Esta es una recopilación de los trabajos sobre la temática que ya existían previo a la década del 90’.En la misma se analizan la actividad enzimática muscular (PFK, fosfofructoquinasa; LDH, lactato dehidrogenasa; SDH, succinato dehidrogenasa; CK, creatinquinasa; PK, proteínquinasa; GS, glucógeno sintetasa; GPh, glucógeno fosforilasa; MDH, malato dehidrogenasa; CS, citrato sintasa; GADPH, gliceraldehído- 3-fosfato dehidrogenasa) y el rendimiento en los sprints repetidos en hombres.
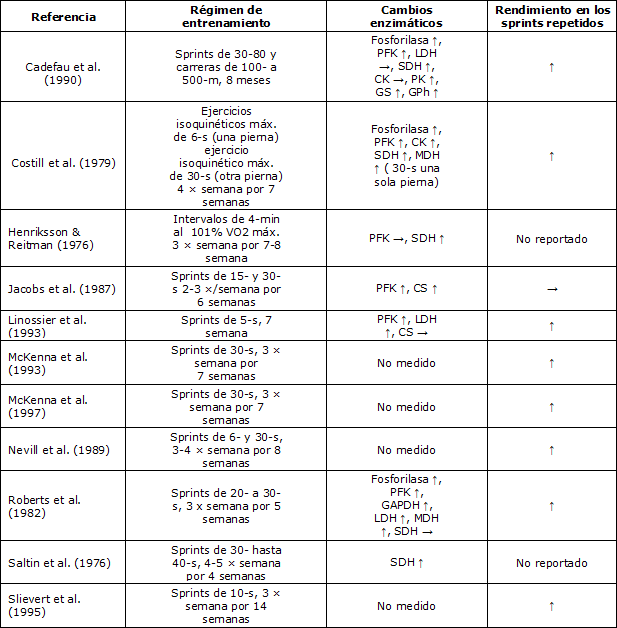 Tabla 1. Adaptaciones aeróbicas y del rendimiento de sprints producidas por entrenamientos de sprints repetidos. ↑ Aumentó, → no se modificó (Mac Dougall et al., 2004).
Tabla 1. Adaptaciones aeróbicas y del rendimiento de sprints producidas por entrenamientos de sprints repetidos. ↑ Aumentó, → no se modificó (Mac Dougall et al., 2004).
A continuación se citarán, con una breve descripción, otros estudios más recientes que continúan en la misma dirección.
Mac Dougall et al. (1998) estudiaron a 22 hombres sanos de 22 ± años edad. Entrenaron con sprints intervalados 3 sesiones semanales durante 7 semanas sobre una cinta ergométrica. El entrenamiento consistió en sprints de 30 segundos (protocolo de Wingate), con 2 a 4 minutos de pausa. El programa comenzó con 4 intervalos con pausas de 4 minutos por sesión en la semana 1 y progresó a 10 intervalos con 2.5 minutos de recuperación en la semana 7. Se tomaron biopsias musculares sobre el vasto lateral del muslo antes y después del programa de entrenamiento.
El programa de entrenamiento resultó en incrementos significativos en la producción de potencia pico (P < 0.05), trabajo total en 30 segundos (P < 0.05), y VO2máx (P < 0.05).
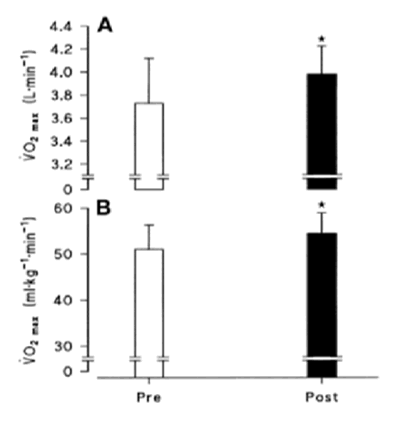 Figura 1. Máximo consumo de oxígeno (VO2máx) absoluto, antes (Pre) y después (Post) del entrenamiento de sprints repetidos. Gráfico B: máximo consumo de oxígeno (VO2máx) relativo, antes (Pre) y después (Post) del entrenamiento de sprints intervalados. Los valores están expresados en unidades absolutas (l/min) y relativas a la masa corporal (mL·kg-1·min-1) en B. * P < 0.05 (Mc Dougall et al. 2004).
Figura 1. Máximo consumo de oxígeno (VO2máx) absoluto, antes (Pre) y después (Post) del entrenamiento de sprints repetidos. Gráfico B: máximo consumo de oxígeno (VO2máx) relativo, antes (Pre) y después (Post) del entrenamiento de sprints intervalados. Los valores están expresados en unidades absolutas (l/min) y relativas a la masa corporal (mL·kg-1·min-1) en B. * P < 0.05 (Mc Dougall et al. 2004).
La actividad enzimática máxima de la hexoquinasa, fosfofructoquinasa, citrato sintetasa, succinato dehidrogenasa, y malato dehidrogenasa también aumentó significativamente después del entrenamiento (P < 0.05).
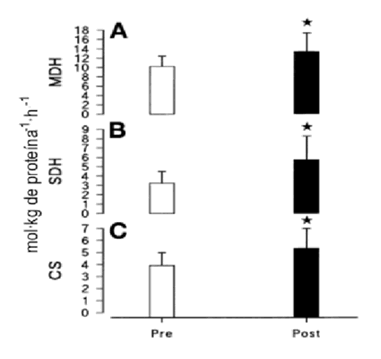 Figura 2. Actividad enzimática máxima para la malato dehidrogenasa (MDH; A), succinato dehidrogenasa (SDH; B), y citrato sintetasa (CS; C), antes y después del entrenamiento de sprints intervalados.* P < 0.05 (Mc Dougall et al., 2004).
Figura 2. Actividad enzimática máxima para la malato dehidrogenasa (MDH; A), succinato dehidrogenasa (SDH; B), y citrato sintetasa (CS; C), antes y después del entrenamiento de sprints intervalados.* P < 0.05 (Mc Dougall et al., 2004).
Stepto et al. (1999) compararon las adaptaciones en la resistencia (valorada con una prueba de ciclismo de 40 km) que producirían distintos protocolos de entrenamientos fraccionados. Para el estudio fueron evaluados 20 ciclistas de resistencia con un consumo de oxígeno máximo pico promedio de 4.8 ± 0.6 L·min-1. Los mismos fueron asignados a 5 grupos que entrenaron con diferentes protocolos, completando 6sesiones durante 3 semanas. Los ciclistas reemplazaron un 15% de su entrenamiento tradicional de resistencia por el entrenamiento fraccionado específico que debía realizar su grupo. Los entrenamientos designados para cada grupo fueron los siguientes (tabla 2):
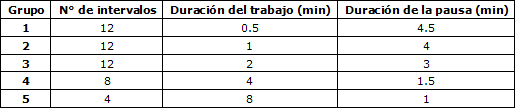 Tabla 2. Entrenamientos intervalados asignados a cada grupo.
Tabla 2. Entrenamientos intervalados asignados a cada grupo.
Los 2 tipos de entrenamientos que produjeron un mayor aumento en la capacidad de resistencia fueron el más específico en cuanto a la intensidad de carrera de competencia (intervalos de 4 minutos al 85% del pico máximo del consumo de O2, 2.8%, 95% CI = 4.3-1.3%) y el de intervalos más cortos y de mayores intensidades (intervalos de 30 s a intensidades del 175% del pico máximo del consumo de O2, 2.4%, 95% CI = 4.0-0.7%). El porcentaje de mejoría de la prueba fue modelado por una función polinómica, procedimiento validado por simulación. Por supuesto que la muestra y el protocolo de evaluación de la resistencia no se corresponde con la especificidad de los deportes de equipo de dinámica intermitente, pero en esta sección se busca principalmente indicar científicamente la relación que existiría entra las adaptaciones aeróbicas y la alta intensidad.
Rodas et al. (2000) estudiaron las adaptaciones aeróbicas producidas por un protocolo fraccionado corto de máxima intensidad. Se estudiaron 5 jóvenes voluntarios activos por 2 semanas donde entrenaron diariamente en un cicloergómetro. El protocolo consistió en periodos de trabajo máximo de 15 s por 45 s de pausa, más períodos de 30 s de esfuerzo máximo por 12 min de pausa. La cantidad de repeticiones aumentaron gradualmente hasta llegar a 7.Se realizaron biopsias musculares antes y después del periodo de entrenamiento sobre el vasto lateral.
Los resultados de las biopsias arrojaron incrementos significativos en la fosfocreatina (31%, P < 0.05) y el glucógeno muscular (32%, P < 0.05). También aumentó significativamente la actividad de enzimas anaeróbicas como la creatinquinasa (44%, P < 0.01), fosfofructoquinasa (106%, P < 0.01) y lactato deshidrogenasa (45%, P < 0.01). Pero al mismo tiempo, se encontraron diferencias significativas en enzimas aeróbicas como la3-hidroxiacil-CoA-deshidrogenasa (60%, P < 0.05) y la citrato sintasa (38%, P < 0.05). Inclusive el VO2máx promedio aumentó de 57.3 (2.6) mL·kg-1·min-1 a 63.8 (3.0) mL·kg-1·min-1 (P < 0.05) en este corto período de 2 semanas, aumentando la máxima carga de trabajo de 300 W (11) a 330 W (21) (P < 0.05); estos cambios también fueron estadísticamente significativos.
Burgomaster et al. (2005) analizaron las adaptaciones aeróbicas que generaría un protocolo de sprints intervalados. El estudio era experimental con un grupo que se sometió al protocolo experimental y un grupo control. El grupo que entrenó consistía en 8 sujetos recreacionalmente activos con una edad promedio de 22 ± 1 años y un VO2pico medio de 45.3 mL·kg-1·min-1. El estudio duró 2 semanas, concluyendo un total de sólo 6 sesiones de entrenamiento separados por 1 o 2 días de recuperación.Cada entrenamiento se realizó en cicloergómetro y consistió en una cantidad de 4 a 7 sprints de 30 s por 4 minutos de recuperación entre los mismos. Antes y después de las 2 semanas de entrenamiento se realizó una prueba para establecer el VO2pico, otra de resistencia que consistía en pedalear hasta la fatiga en un cicloergómetro electrónicamente frenado en donde se buscó que la carga fisiológica represente un 80% del VO2pico, y biopsias musculares sobre la pierna.
Se encontraron diferencias significativas (P < 0.05) en la capacidad de sprint medida a través de la potencia (W) de los mismos.
La misma tendencia se observó en la máximaactividad de la enzima aeróbica citrato sintasa (CS), aumentando en un 38% post entrenamiento (P < 0.05).
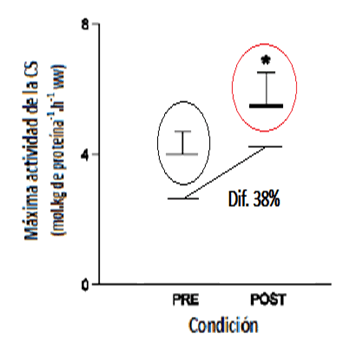 Figura 3. Variación de la máxima actividad de la enzima citrato sintasa antes y después del período de entrenamiento de sprints intervalados. * P < 0.05 (modificado de Burgomaster et al., 2005).
Figura 3. Variación de la máxima actividad de la enzima citrato sintasa antes y después del período de entrenamiento de sprints intervalados. * P < 0.05 (modificado de Burgomaster et al., 2005).
El contenido de glucógeno muscular también se encontró aumentado en un 26% luego del período de 2 semanas (P < 0.05).
En cuanto a la capacidad de resistencia global, medida con el test de resistencia en cicloergómetro, siguiendo con la misma línea se halló incrementada en un 100% (551 VS 265 min, P < 0.05).
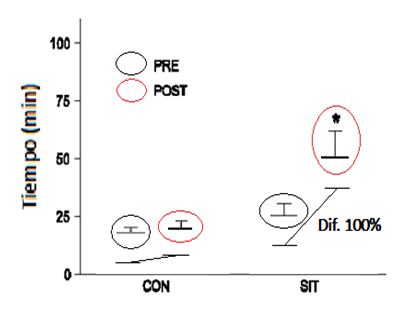 Figura 4. Minutos recorridos en una prueba de resistencia hasta la fatiga al 80% del VO2pico antes y después del período de entrenamiento para el grupo control y el grupo SIT. * P < 0.05 (modificado de Burgomaster et al., 2005).
Figura 4. Minutos recorridos en una prueba de resistencia hasta la fatiga al 80% del VO2pico antes y después del período de entrenamiento para el grupo control y el grupo SIT. * P < 0.05 (modificado de Burgomaster et al., 2005).
En el estudio se concluye que el entrenamiento corto e intenso (~15 min de ejercicio fraccionado intenso por 2 semanas) de sprints intervaladosincrementa el potencial oxidativo del músculo y la capacidad de resistencia durante una prueba aeróbica intensa de ciclismo, en sujetos recreacionalmente activos.
Burgomaster et al. (2007) estudiaron las adaptaciones aeróbicas producidas por un entrenamiento de sprints intervalados,tomando como variables el contenido de proteínas relacionas con el transporte de glucosa, de lactato y de ácidos grasos. La muestra estaba constituida por 8 sujetos masculinos activos, con una edad promedio de 22 ± 1 años y un VO2pico medio de 50.2 mL·kg-1·min-1. Entrenaron 3 sesiones semanales durante 6 semanas, con un protocolo en cicloergómetro de 4 a 6 sprint de 30 s por 4 min de pausa. Se hicieron biopsias musculares sobre el vasto lateral antes y luego de la primera y sexta semana de entrenamiento. También 1y 6 semanas después de finalizado el entrenamiento.
La capacidad oxidativa del músculo, reflejada en el contenido proteico del citocromo C oxidasa subunidad 4 (COX4) de la cadena transportadora de electrones mitocondrial, se encontró aumentada en un 35% luego de la primer semana de entrenamiento por sprints, y se mantuvo significativamente elevada inclusive luego de 6 semanas de desentrenamiento (P < 0.05).
La proteína relacionada con el transporte de glucosa (GLUT4) en el músculo se incrementó también luego de la primer semana de entrenamiento y se mantuvo elevada en relación al valor inicial luego de 6 semanas de haber finalizado el entrenamiento (P < 0.05).
En relación al transporte del lactato, se observaron incrementos significativos (P < 0.05) de los transportadores de monocarboxilato 4 (MCT4)luego de la primera y sexta semana de entrenamiento. Los transportadores de monocarboxilato 1 (MCT1) se encontraron aumentados luego de la sexta semana de entrenamiento en relación a los valores pre-entrenamiento, y se mantuvieron elevadosluego de la primer semana post entrenamiento (P < 0.05). Paralelamente a estos incrementos se sumó el dato de que no se apreciaron variaciones sobre las traslocasas de los ácidos grasos (AG) asociadas con la ligadura proteica de los AG (FABPpm) (P < 0.05).
Los autores concluyeron que el entrenamiento corto de máxima intensidad induce a rápidas adaptaciones en la capacidad oxidativa del músculo esquelético, pero con divergentes efectos sobre la capacidad de las proteínas asociadas al transporte de glucosa, lactato y ácidos grasos.
Little et al. (2010) analizaron las adaptaciones aeróbicas producidas por un entrenamiento intermitente de alta intensidad (según la caracterización propuesta se encontraría justo en el límite entre una metodología intervalada y una intermitente, por lo cual podria encuadrar dentro de cualquiera de las 2 definiciones), tomando como variables la actividad de la enzima aeróbica citrato sintetasa, el contenido proteico de GLUT4, el contenido proteico de los citocromos C oxidasa de la cadena transportadora de electrones mitocondrial subunidad 2 y 4 (COX2 y COX4), el factor A de transcripción mitocondrial, y el contenido total SIIRT1, propuesto como activador del marcador mitocondrial del metabolismo de los carbohidratos PGC-1α (coactivador-1α de receptor-γ activado por proliferadores de peroxisomas) y de la biogénesis mitocondrial. La muestra estuvo constituida por 7 hombres jóvenes activos (realizaban actividad física 2 a 3 veces semanales) de 21 ± 0.4 años de edad y un VO2pico de 46 2 mL·kg-1·min-1. El protocolo propuesto fue de 6 entrenamientos divididos en 2 semanas, donde realizaban de 8 a 12 intervalos de 60 s, al ~100% del pico máximo de potencia extraído del test de rampapara medir el VO2pico (355 ± 10W),por 75 s de pausa entre los esfuerzos. Se realizaron biopsias sobre el vasto lateral de la pierna para extraer las muestras.
El entrenamiento aumentó la máxima actividad y el contenido proteico de la citrato sientetasa en un ~16 y 20% (P = 0.01). El contenido proteico de los citocromos C oxidasa aumentó en un 35% para el COX2 (P = 0.01) y en un 38% para el COX4 (P = 0.002). El contenido proteico del GLUT4 y el contenido de glucógeno muscular también se encontraron incrementados en un 119% (P = 0.04) y un 17% (P = 0.05) respectivamente. El factor A de transcripción mitocondrial también se encontró significativamente aumentado (P < 0.05). El contenido total SIIRT1 también se halló significativamente incrementado luego del proceso de entrenamiento (56%, P < 0.05).
En el estudio se concluye que un modelo de bajo volumen y alta intensidad es un estímulo potente para aumentar la capacidad mitocondrial del músculo esquelético y mejorar el rendimiento físico.
da Silva et al. (2010) analizaron la relación entre el rendimiento en una prueba de sprints repetidos y distintas variables de la condición aeróbica como los son el VO2máx, la vVO2máx (velocidad de VO2máx o VAM) y la velocidad de inicio de acumulación de lactato en sangre (vOBLA). En el estudio participaron 29 jugadores de fútbol de elite brasileros de primera división de 2 niveles de competencia nacional (liga A y B).La edad promedio era de 17.9 ± 1.0 años. Se realizaron 2 protocolos de evaluación, un test incremental para medir el VO2máx y la vOBLA y un test para medir la RSA que consistió en 7 sprints de 34.2 m con 25 s de pausa entre los mismos.
Se encontraron significativas correlaciones negativas entre el tiempo medio (MT) de la prueba de RSA con la vOBLA y la vVO2máx (r = -0.49, P < 0.01; r = -0.38, P < 0.05, respectivamente). También se encontraron significativas correlaciones negativas entre el decrecimiento de la capacidad de sprint (Sdec) y la vOBLA (r = -0.54), la vVO2máx (r = -0.49) y el VO2máx (r = -0.39).
El estudio indica que los sprints repetidos tienen relación con la capacidad aeróbica, este hecho se evidencia en las significativas correlaciones entre distintas variables de esta capacidad y la habilidad de repetir sprints (RSA). Las relaciones no se encuentran muy elevadas porque como es sabido existen otros factores de marcada importancia que influyen en esta capacidad, como lo son los sistemas anaeróbicos de re síntesis de ATP. Pero de que el sistema aeróbico también realiza su aporte parecería ser un hecho en la actualidad.
Evidentemente el sistema aeróbico también contribuye energéticamente para la ejecución de esfuerzos repetidos de máxima intensidad considerados tradicionalmente como ejercicios puramente anaeróbicos. Esto da sentido a los resultados reflejados en los estudios presentados anteriormente donde las adaptaciones del sistema aeróbico generadas por distintos protocolos de sprints intervalados y repetidos comprueban la implicancia del sistema oxidativo, y por consiguiente su estímulo y posterior adaptación, en esta modalidad de ejercicios intensos.
Una de las posibles explicaciones fisiológicas a esta relación se basa en el mecanismo denominado como lanzadera de la fosfocreatina (“shuttle de la PCr”), donde diversos estudios han establecido la directa relación que existiría entre el sistema anaeróbico de los fosfágenos y el sistema oxidativo (Besseman & Geiger 1981; Barbours et al., 1984; Yoshizaki et al., 1987; Savabi et al., 1988; Schnyder et al., 1991; Wallimann et al., 1992; Andrienko et al., 2003; Vendelin et al., 2004; Brooks et. al., 2005;Freyssenet, 2007). Entonces el sistema oxidativo resultaría relevante en la posibilidad de resintetizar permanentemente el grupo fosfato promovido por la PCr. Este es sólo un ejemplo donde se puede divisar una relación directa entre estos sistemas. Hay que mencionar que este mecanismo se produce en el cuerpo en todo momento, incluso en reposo. Por lo tanto la discusión podría establecerse en torno al papel que jugaría este mecanismo en esfuerzos intensos, incluso maximales, donde se utiliza en una alta magnitud las reservas de PCr, en forma sistemática en el caso de que las intensidades elevadas y/o máximas se repitan sucesivamente en el tiempo. Por ende la demanda de restitución de la misma (PCr) por parte de la mitocondria (relación entre mitocondria y citosol) se podría ver incrementada, por consiguiente también el estimulo adaptativo sobre esta organela celular. Sin dudas que hablamos solamente de una hipótesis, una interpretación fisiológica más para la discusión.
Existe una amplia cantidad de estudios científicos como los presentados que indicarían esta relación entre la alta intensidad y las adaptaciones aeróbicas, como el de Tabata et al. (1996) o el de Harmer et al. (2000) donde concluyen que el entrenamiento de sprints repetidos resulta en una reducción de la generación anaeróbica de ATP durante el ejercicio intenso, lo que sugeriría que el aporte aeróbico sería realzado, dando lugar al aumento del tiempo hasta la fatiga.
Distintas revisiones bibliográficas también abordarían el tema desde esta línea científica donde parece ser que los distintos tipos de entrenamientos fraccionados cortos de alta y máxima intensidad producirían importantes y significativas adaptaciones aeróbicas, indicando que existiría una relación entre la elevada intensidad en protocolos de esfuerzos repetidos y la capacidad y potencia aeróbica (Finn, 2001; Coyle, 2005; Spencer et al., 2005; Barbero Álvarez et al., 2006a y b).
En los esfuerzos de RSA el sistema oxidativo aportaría energía desde el primer sprint, al igual que el sistema de fosfágenos y la glucólisis rápida, estos últimos en magnitudes superiores por supuesto (Spencer et al., 2005) y a medida que se suceden los mismos el aporte del sistema de fosfágenos y de la glucólisis rápida caerían. En este sentido Gaitanos et al. (1993), en un protocolo de 10 sprints de 6 s, reportó que el aporte estimado de la PCr para resintetizar el ATP durante el primer sprint fue de 49,6% y el de la glucólisis rápida del 44,1%, pero a raíz de la inhibición producida en mayormedida sobre la glucólisis rápida a medida que se suceden los sprints, en el décimo la contribución de la PCr fue de un 80,1%, mientras que el de la glucólisis fue de tan sólo un 16,1%. En el décimo sprints la glucogenólisis cayó 11 veces y la glucólisis 8 veces, aunque la degradación del glucógeno total había caído un 37%. Sin embargo, en términos absolutos la resíntesis de ATP a partir de la PCr en el último sprints cayó en un 42,8%, mientras que la producida a partir de la glucólisis rápida descendió en un 87%. Este fenómeno fisiológico parece venir acompañado de un incremento en el aporte del sistema oxidativo en la medida que se suceden los sprints repetidos con pausas incompletas (Spriet et al., 1989; Gaitanos et al., 1993; Bogdanis etal., 1996; Chritmass et al., 1999; citados por Barberos Álvarez et al., 2006a y b, p. 3). El aporte total del sistema oxidativo en un esfuerzo de sprints repetidos sería de entre el 28 y el 40%, tanto en protocolos desprints más largos (30 s) como más cortos (inferiores a 22 s) (Medbø & Tabata, 1989; Withers et al., 1991; Medbø et al., 1999; Spencer & Gastin, 2001; citados por Spencer et al., 2005, p. 1031).También se ha indicado un aumento en el VO2máx mientras se suceden los sprints en esfuerzos de estas características (Hamilton et al., 1991; Tomlin & Wenger, 2001; citados por Barberos Álvarez et al. 2006a y b, p. 7; ), lo cual guardaría una directa relación con el aumento del aporte del sistema oxidativo anteriormente mencionado. No obstante, son necesarios más estudios que permitan determinar con mayor precisión el momento de participación y el tipo de contribución del metabolismo oxidativo durante un esfuerzo de RSA.
ADAPTACIONES AERÓBICAS: CONTINUOS VS. SPRINTS INTERVALADOS
Gibala et al. (2006) compararon las adaptaciones aeróbicas generadas por un protocolo continuo de entrenamiento de la resistencia (protocolo continuo extensivo) con las producidas por un entrenamiento de intensidad máxima y bajo volumen (protocolo de sprints intervalados). En el estudio participaron 16 sujetos masculinos activosde 21 ± 1 años en promedio con un VO2pico medio de 4.0 ±0.21 L.min-1. Se formaron 2 grupos de 8 sujetos, uno realizó un entrenamiento de sprints intervalados, «Sprint interval training» (SIT), y el otro grupo desarrolló un entrenamiento tradicional de resistencia, “Endurance Training” (ET).Ambos grupos entrenaron en 6 ocasiones en el transcurso de 2 semanas. El grupo SIT por sesión de entrenamiento ejecutó entre 4 y 6 esfuerzos de máxima intensidad de 30 s en cicloergómetro (intensidad del 250% del VO2pico) por 4 min de recuperación entre los períodos de trabajo. El grupo ET se entrenó también en cicloergómetro durante 90 a 120 min a una intensidad del 65% del VO2pico durante cada sesión de entrenamiento.Se realizaron biopsias musculares, en la porción lateral del muslo, antes y después del período de entrenamiento.
Las adaptaciones aeróbicas fueron similares en ambos grupos, reflejadas en un similar aumento de la máxima actividad del citocromo C oxidasa (COX) y el contenido proteico del COX subunidad 2 y 4 (principales efectos, P ≤ 0.05). Cabe recordarque estos citocromos son base fundamental del funcionamiento de la cadena transportadora de electrones donde se produce la fosforilación oxidativa, generadora de la mayor parte del ATP que sintetiza el sistema aeróbico.
También fueron similares los efectos sobre el aumento en la capacidad buffer y el contenido de glucógeno muscular (principales efectos, P ≤ 0.05).
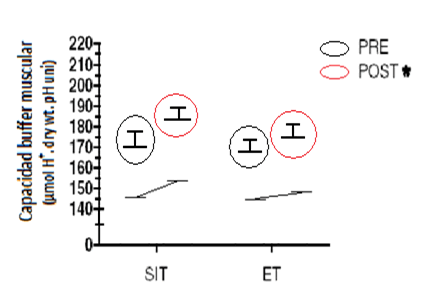 Figura 7. Capacidad buffer muscular antes y después del período de entrenamiento para el grupo SIT y ET. * P ≤ 0.05 (modificado de Gibala et al., 2006).
Figura 7. Capacidad buffer muscular antes y después del período de entrenamiento para el grupo SIT y ET. * P ≤ 0.05 (modificado de Gibala et al., 2006).
El estudio concluye que si se considera la gran diferencia en el volumen de entrenamiento, estos datos demuestran que el SIT (entrenamiento intervalado de sprint) es una estrategiamás eficiente para inducir rápidas adaptaciones en el músculo esquelético y en el rendimiento del ejercicio en comparación con el ET (entrenamiento continuo de resistencia) en hombres jóvenes activos.
El tiempo total de entrenamiento realizado por el grupo SIT fue de 2 horas y media, mientras que el tiempo total del grupo ET fue de 10 horas y media, lo que implica que el volumen de entrenamiento del grupo SIT fue un 90% inferior al del grupo ET (630 VS. 6500 KJ).
Burgomaster et al. (2008) retomaron la comparación entre el entrenamiento corto e intenso y el de menores intensidades con altos volúmenes de dinámica o característica continua. En el estudio participaron 10 hombres y 10 mujeres, que se distribuyeron 5 hombres y 5 mujeres para el grupo que entrenó con sprints intervalados (SIT) y la misma cantidad para el que entrenó con una metodología continua (ET). Los sujetos eran considerados activos pero desentrenados, con una edad promedio de 23 ± 1 años. El entrenamiento SIT consistió en 6 repeticiones de 30 s de intensidad máxima en cicloergómetro por 4.5 minutos de pausa, 3 días por semana. El entrenamiento ET consistió en entrenar 5 días semanales en forma continua durante 40 a 60 min a una intensidad del 65% del VO2pico. Ambos grupos se entrenaron durante 6 semanas. Se realizaron biopsias musculares sobre el vasto lateral del muslo antes y después del programa de entrenamiento.
Ambos protocolos indujeron similares adaptaciones (P < 0.05) en los marcadores mitocondriales del metabolismo de los carbohidrato, la oxidación de lípidos (máxima actividad de la 3-hidroxiacil-CoA-deshidrogenasa) y el contenido proteico del coactivador-1α de receptor-γ activado por proliferadores de peroxisomas. Luego del entrenamiento se observó una reducción en la utilización de fosfocreatina y glucógeno durante el ejercicio y se calculó que la tasa de oxidación corporal total de carbohidratos y lípidos disminuyó y aumentó respectivamente (efectos principales, P < 0.05).
En el estudio se concluye que dada la gran diferencia en el volumen de entrenamiento entre ambos grupos (1.5 contra 4.5 horas semanales), el entrenamiento a intervalos de máxima intensidad es una estrategia más eficiente en relación al tiempo para generar rápidas adaptaciones en la capacidad oxidativa del músculo esquelético, induciendo también específicas adaptaciones durante el ejercicio, en comparación con el entrenamiento continuo de resistencia.
Estos estudios y otros, como el de Kohan et al. (2008), parecen indicar que la resistencia y sus bases aeróbicas no están ligadas única y necesariamente a métodos de bajas y moderadas intensidades, ya que los métodos de mayores intensidades, incluso máximas, producen también adaptaciones aeróbicas en grados similares, e incluso mayores, como los estudios citados lo han señalado. Hay que recordar y reconocer la inmensa diferencia entre los volúmenes utilizados para cada modalidad metodológica en estos estudios. A futuro podría ser importante el comparar los resultados con un aumento en el volumen del entrenamiento fraccionado intenso. Cabe mencionar que un manejo direccionado específico en el fraccionamiento de la carga, en relación al tiempo de trabajo y los tiempos de las micro y macro pausas, permite aumentar estos volúmenes de esfuerzos de elevadas y máximas intensidades.
Por otro lado dentro del contexto de los deportes de equipo, es digno de destacar que los entrenamientos más específicos y de mayores intensidades propondrían un estímulo adaptativo fibrilar (neuromuscular) de mayor especificidad y magnitud (Greig et al., 2006), al mismo tiempo que también producirían importantes adaptaciones anaeróbicas, como aumento de la actividad de enzimas anaeróbicas (Mac Dougall et al., 1998), en transportadores de monocarboxilato (Burgomaster et al., 2007) y en el contenido muscular de PCr (Burgomaster et al., 2008), entre otras. Este hecho las podría colocar como elecciones más adecuadas desde una perspectiva más amplia e integral.
ADAPTACIONES AERÓBICAS: INTERVALADO VS. SPRINTS REPETIDOS (RSA)
Ferrari Bravo et al. (2008) compararon las adaptaciones aeróbicas producidas por un protocolo de entrenamiento intervalado con las generadas por un entrenamiento de sprints repetidos (RSA). La muestra consistió en 42 jugadores de fútbol, 22 profesionales junior (17.3 ± 0.6 años, 71 ± 5.6 kg, 179.3 ± 4.8 cm y 9.3 ± 2.7% de masa grasa estimada) y 20 jugadores de primera división amateurs (24.3 ± 5.4 años, 76 ± 5.4 kg, 179.4 ± 4.8 cm y 11.0 ± 3.8% de masa grasa estimada).El grupo ITG (“interval training Group”), N = 21, entrenó con 4 carreras de 4 min al 90-95% de la FCmáx, con 3 min de recuperación activa entre los esfuerzos (60-70% de la FCmáx). La frecuencias cardíacas eran controladas en forma directa con sistemas de telemetría (Vantage NV, XTrainer, modelos S610 y S710, Kempele, Polar, Finlandia). El grupo RSG (“Repeated Sprint Group”), N = 21, entrenó con 3 series de 6 sprints de 40 m (7 s de esfuerzo máximo aproximadamente por sprint), con 20 s de pausa pasiva entre los mismos y 4 min entre serie. Los sprints eran con cambios de direcciones de 180° cada 10 m en las primeras 3 semanas y cada 20 m en las otras 4. Ambos grupos se entrenaron por un total de 7 semanas durante el período competitivo, donde se destinaba una fracción de 2 de los 4 entrenamientos de 90 min que realizan semanalmente para los correspondientes protocolos de entrenamiento. Nunca los entrenamientos fueron en días consecutivos.
Antes del período de entrenamiento y después de las 7 semanas se realizaron las siguientes mediciones: VO2máx, el punto compensatorio de respiración, la resistencia específica del fútbol a través del YOYO de recuperación intermitente (Recovery Test), el tiempo de sprint en 10 m, la altura del salto y la potencia (CMJ y SJ), y la capacidad de repetir sprints (RSA).
El VO2máx post entrenamiento, en ambos casos, aumentó por debajo de un 6%, de 52.8 ± 3.2 a 56.3 ± 3.1mL·kg-1·min-1 para el grupo ITG y de 55.7 ± 2.3 a 58.5 ± 4.1 mL·kg-1·min-1para el grupo RSG. Sin embargo amplias fueron las diferencias en la capacidad de repetir sprints y en la capacidad de resistencia intermitente donde en ambos casos el grupo RSG obtuvo beneficios más elevados que el grupo ITG. Para la prueba de RSA en el grupo ITG no se reportaron mejoras significativas (P = 0.55), mientras que en el grupo RSG el rendimiento en la misma mejoró significativamente en un 2,1% (P = 0.001). En cuanto a la prueba de resistencia intermitente el grupo ITG mejoró el rendimiento en un 12,5% (P < 0.01), mientras que las diferencias en el grupo RSG fueron de un 28,1% (P < 0.001). La capacidad de repetir sprints evaluada antes y después de la pretemporada con el “Repeated-Sprint Ability Shuttle Test” (6 sprints de 40 m, 20 + 20 m, separados por 20 s de pausa entre los esfuerzos) arrojó una disminución del tiempo de sprint medio que va de 7.53 ± 0.21 a 7.37 ± 0.17 s(P = 0.001) para el grupo RSG y de 7.42 ± 0.22 a 7.40 ± 0.22 s (P = 0.55) para el grupo ITG.
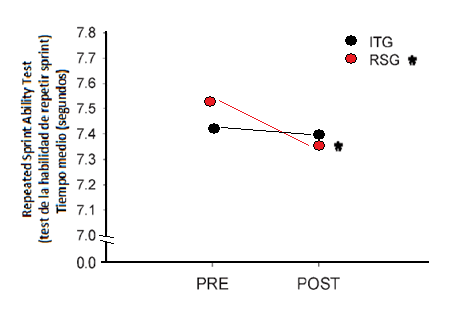 Figura 9. Tiempo medio de los sprints del test de RSA (indicador de la capacidad de repetir sprints) antes y después del período de7 semanas de entrenamiento para el grupo ITG y RSG.* P < 0.001 (modificado de Ferrari Bravo et al., 2008).
Figura 9. Tiempo medio de los sprints del test de RSA (indicador de la capacidad de repetir sprints) antes y después del período de7 semanas de entrenamiento para el grupo ITG y RSG.* P < 0.001 (modificado de Ferrari Bravo et al., 2008).
En cuanto a la resistencia específica del fútbol, como se señaló, la misma fue evaluadaa partir del YOYO de recuperación intermitente,propuesta validada en numerosas ocasiones para cumplir con la función, a partir de la medición de la sensibilidad de la propuesta para diferenciar entre niveles de competencia en comparación con otros tests y de correlaciones con los metros a alta intensidad recorridos durante la competencia (Krustrup & Bangsbo, 2001; Krustrup et al., 2003; Krustrup et al., 2005; Krustrup et al., 2006; Castagna et al., 2005; Castagna et al., 2008; Bangsbo et al., 2008). El protocolo específicamente evalúa la capacidad de ejecutar acciones cortas e intensas (de distintas intensidades, ordenadas protocolarmente en un orden progresivo) con pausas semi-completas propias del rendimiento físico relacionado con la resistencia intermitente específica de este tipo de disciplinas deportivas.
Para el grupo RSG los metros promedios recorridos en la prueba antes y después del período de entrenamiento fueron de 1917 ± 439 y 2455 ± 488 m respectivamente, mientras que para el grupo ITG fueron de 1846 ± 329 y de 2077 ± 300 m respectivamente. Esto indica que quienes entrenaron con sprints repetidos aumentaron un 28.1% (P < 0.001) su capacidad de resistencia intermitente específica contra un aumento del 12.5% (P < 0.01) que obtuvieron quienes entrenaron con el método intervalado.
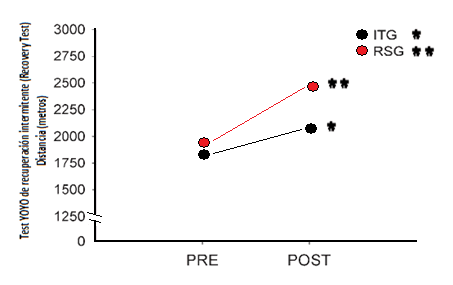 Figura 10. Promedio de metros recorridos en el Recovery Test antes y después del período de7 semanas de entrenamiento para el grupo ITG y RSG. * P < 0.01; ** P < 0.001 (modificado de Ferrari Bravo et al., 2008).
Figura 10. Promedio de metros recorridos en el Recovery Test antes y después del período de7 semanas de entrenamiento para el grupo ITG y RSG. * P < 0.01; ** P < 0.001 (modificado de Ferrari Bravo et al., 2008).
Fernarndez-Fernandez et al. (2012) también compararon un entrenamiento intervalado, con un protocolo con cierto grado de especificidad en relación al deporte en cuestión (tenis), con uno de sprints repetidos. 31 tenistas competitivos (competían a nivel nacional con un ranking nacional entre 150 y 250) se dividieron en 3 grupos, 11 en el grupo intervalado «High Intensity interval training» (HIIT), 12 en el de sprints repetidos «Repeated Sprint Training» (RST) y 9 en el grupo control (CON). La edad promedio era de 22.6 ± 4.8 años (HIIT), de 21.2 ± 5.1 años (RST) y de 22.1 ± 3.3 años (CON). El VO2pico medio era de 56.3 ± 4.0 mL·kg-1·min-1 (HIIT), de 55.6 ± 5.0 mL·kg-1·min-1 (RST) y de 57.7 ± 3.9 mL·kg-1·min-1 (CON). El entrenamiento intervalado consistía en 3 series de 3 repeticiones de 90 s (intensidad de un 90-95% de la FCmáx) con 180 s de pausa activa (intensidad de un 70% de la FCmáx) entre cada repetición. Este entrenamiento fue controlado a partir de los beep de un CD (protocolo «Hit and Turn Test«). El protocolo del grupo que entrenó con sprints repetidos consistió en la replica del «RSA Shuttle Test», 3 series de 10 repeticiones de 22 metros de sprints con 20 s de pausa pasiva entre repetición.Ambos grupos realizaron un game de un juego 2 vs. 1 entre series (la intensidad fue fjada entre el 75% y el 85% de la FCmáx), donde se realizaba la macro pausa entre las mismas que era de 8 min.
Se midió el VO2pico en forma directa con un test incremental en tapiz rodante, la resistencia especifica del tenis a través del «Hit and Turn Tennis Test» y la RSA a través del «Repeated-Sprint Ability Shuttle Test» y la altura alcanzada en saltos CMJ.
El VO2pico se incrementó significativamente tanto en el grupo HIIT (6%, P = 0.008) como en el RST (4,9%, P = 0.010). La resistencia específica también aumentó significativamente en ambos grupos (HIIT 28.9% VS. RST 14.5%, P = 0.05). De los 3 grupos el grupo RST fue el único que manifestó una mejora significativa en el test de RSA, evidenciada en la reducción del tiempo medio de sprint (3.8%, P = 0.05). El grupo control no mostró diferencias significativas en las variables analizadas. No fueron observadas diferencias significativas en los saltos para ningún grupo.
Los autores concluyeron que ambas metodologías inducen similares adaptaciones sobre la capacidad aeróbica, pero que el entrenamiento de sprints repetidos resulta ser una estrategia más eficaz ya que produce adaptaciones aeróbica específicas al mismo tiempo que mejora la RSA.
EL LACTATO (La–), LA ALTA INTENSIDAD Y LA BIOGÉNESIS MITOCONDRIAL
Antes de ingresar en lo específico de la sección se profundizará sobre la relación entre el lactato y la acidosis metabólica ya que el tema se encuentra en una profunda discusión (Prakash et al., 2008) y para que lo que se presente no se preste para confusas interpretaciones se buscará describir una interpretación científica que sirva como base al tema en cuestión.
Desde la interpretación de los principios físico-químicos de la estructura del lactato, tomando el pKa del ácido láctico (3,87 a 37 °C), al pH de la célula muscular (6.2 – 7.0) difícilmente en la misma se podría generar esta sustancia como se sostiene en gran parte de la bibliografía en relación al tema. Lo que se formaría es la base conjugada de este ácido (su anión), el lactato (La–), que tiene características diferentes al mismo, partiendo de la base de no ser un ácido, por lo tanto no sería propenso a generar acidez liberando H+ al medio. Desde esta interpretación la glucólisis generaría la forma ionizada del ácido pirúvico (piruvato) y por lo tanto consecuentemente lo que se formaría a partir del mismo es la forma ionizada del ácido láctico (lactato) (Martin et al., 2001; Robergs, 2001; Robergs et al, 2004; Mendoza Medellin A., 2008). Por esto lo que la glucólisis rápida generaría como producto final es el lactato y no el ácido láctico. Entonces el lactato no sería el causante de la acumulación de H+ en el citosol y por lo tanto de la acidez muscular, pero sí lo podria ser la glucólisis rápida ya que la alta intensidad relacionada con el uso predominante de la misma provocaría una alta tasa de hidrólisis de ATP lo cual generaría una alta concentración de iones H+. También la fase preparatoria de la glucólisis utiliza ATP, lo cual incrementaría el contenido de H+ (Robergs et al., 2004). Cabe destacar que el tema de la acidosis metabólica aún hoy se encuentra en constante discusión (Prakash et al., 2008).
Entonces la elevada producción de La– que generalmente producen las metodologías de mayores intensidades ayudaría a estimular el sistema de producción del mismo, lo cuál sería beneficioso ya que ese proceso de formación de La– permite mantener el potencial redox citosólico (NAD+/NADH), apoya el flujo continuo de sustratos a través de la fase dos de la glucólisis y de este modo permite continuar la regeneración de ATP a partir de la glucólisis. Paralelamente por cada molécula de piruvato catalizada a lactato y NAD+, se consumiría un H+, lo cual convierte a la reacción catalizada por la LDH en un eficiente mecanismo de control de la acidez celular debido al consumo sistemático de protones que dicha reacción produce (Robergs, 2001; Robergs et al, 2004; Mendoza Medellin, 2008; Prakash et al, 2008). Por esta razón los organismos capaces de generar altas concentraciones de lactato, como de removerlo y reutilizarlo rápidamente, poseerían la capacidad de perpetuar por lapsos más extensos los esfuerzos intensos.
Entonces desde esta interpretación parecería que la acidez muscular (acumulación de H+ en el citosol de la célula fibrilar), que algunos estudios relacionarían con posibles deterioros y mermas de la contracción muscular, podría generarse por otros mecanismos como la hidrólisis del ATP, aumentada por la alta intensidad relacionada con la predominancia en la utilización de la glucólisis rápida y por su fase preparatoria, ya que por cada rompimiento de ATP se liberaría un H+ al medio (Robergs, 2001; Robergs et al, 2004; Mendoza Medellin, 2008; Prakash et al., 2008) y/o a través de otros mecanismos como lo proponen diversas teorías como lo es la basada en el principio de electroneutralidad, que señala la posibilidad de que la acidez muscular se genere por causa de un desequilibrio de cargas que generarían algunas sustancias, como el La– (Mendoza Medellin, A., 2008). Esta última se basa en principios fisicoquímicos propuestos por el modelo de Stewart (Esper R.C., et al, 2008). En este caso el agua celular (H2O) podría ser una molécula que, por motivo de estas diferencias de cargas con otras moléculas, podría liberar H+ al medio induciendo a un descenso del pH en la fibra muscular.
Ingresando sí ahora en lo específico del tema del lactato, la alta intensidad y las beneficiosas adaptaciones aeróbicas que produciría la elevada producción de La– (como la biogénesis mitocondrial),es que se señalará que la alta intensidad no sólo estimularía el determinante mecanismo de producción del La–, sino que también los sistemas de remoción de La– y H+ como el de los transportadores de monocarboxilatos (MCT) (Baker et al., 1998; Pilegaard et al., 1999;Juel et al., 2003; Bickham et al., 2006; Edge et al., 2006; Green, 2007; Thomas et al., 2007; Messonnier et al., 2007; Huang et al., 2007; Burgomaster et al., 2007). Este mecanismo es denominado “Shuttle del La–”. Por lo tanto con las metodologías fraccionadas de alta intensidad, que pongan en funcionamiento a alta tasa el sistema de producción y remoción del La– (“lactate turnover”) y los sistemas de control de la acidez muscular, se desarrollarían los mismos repercutiendo favorablemente en la capacidad de resistencia física, en especial en el marco de los deportes de equipo donde en ocasiones estos sistemas son sobrecargados por las características generales del esfuerzo que se realiza durante la competencia.
La alta producción del lactato estimularía también el ciclo de Krebs y la cadena transportadora de electrones ya que parte del mismo se oxida dentro de la mitocondria a través de estos mecanismos fisiológicos, lo que se denomina fisiológicamente como lanzadera intra-celular del lactato (“Intracelular Lactate Shuttle”) (Dubouchaud et al., 2000; Gladden et al., 2004). Al mismo tiempo este La– activaría vías de señalización de genes que estimularían la biogénesis mitocondrial (Hashimoto et al., 2007). Por lo tanto la alta intensidad, el lactato y las adaptaciones aeróbicas tendrían una directa relación encadenada, ya que la primera generaría una alta tasa de producción de La–, y este diversos mecanismos fisiológicos y adaptaciones, muchas consideradas aeróbicas como la biogénesis mitocondrial, como las anteriormente señaladas.
CAPILARIZACIÓN FIBRILAR Y ALTA INTENSIDAD
Jensen et al. (2004) plantean que esfuerzos de mayores intensidades estimularían en mayor medida la capilarización fibrilar que los de menores. Estos autores realizaron 2 estudios, en el primero participaron 6 sujetos sanos de una edad promedio de 25.3 ± 1.2 años, 82.8 ± 4.8 kg y un VO2máx de 50.2 ± 0.5 mL·kg-1·min-1. Para el segundo estudio participaron 7 sujetos desentrenados de una edad promedio de 23.9 ± 1.4 años, 76.1 ± 2.7 kg y un consumo máximo de O2 de 45.4 ± 1.9 mL.min-1 .kg-1. Se utilizaron 2 protocolos de ejercicio fraccionado de extensiones de rodilla. Para el estudio 1 el protocolo de entrenamiento consistió en un trabajo de 1 min al 150% del VO2máx por 3 min de pausa durante 1 hora. Para el estudio 2 el entrenamiento consistió en realizar las extensiones de pierna al 90% del VO2máx durante 1 min de trabajo por 30 s de pausa. El tiempo total de cada entrenamiento fue de 1 hora. En ambos estudios se entrenó durante 6.8 ± 0.3 semanas, durante las 2 primeras se realizaron 3 sesiones semanales, durante las 2 siguientes 4 sesiones, y durante las últimas 3 eran 5 sesiones por semana. Se realizaron biopsias musculares antes del periodo de entrenamiento y a las 2, 4 y 6 semanas, en la pierna entrenada y en la no entrenada.
Luego de 4 semanas se observaron diferencias significativas (P < 0.05) en el radio de capilarización por fibra para el estudio que entrenó con mayor intensidad (estudio 1: 2.4 ± 0.1 vs 1.7 ± 0.1) y el número de células Ki-67 positivas que indican la proliferación capilar (estudio 1: 0.18 ± 0.05 vs 0.00 ± 0.01). Ni la posición de la proliferación de las células endoteliales ni la capilarización se relacionó con el tipo de fibras musculares.
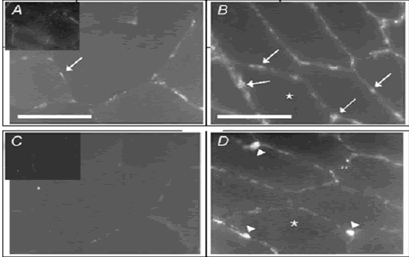 Figura 11. Capilares CD31-positivos antes (A) y después de 4 semanas de entrenamiento fraccionado intenso y proliferación de células Ki-64 positivas (indicando desarrollo de capilares) antes (C) y después (D) de 4 semanas de entrenamiento intenso (Jensen et al., 2004).
Figura 11. Capilares CD31-positivos antes (A) y después de 4 semanas de entrenamiento fraccionado intenso y proliferación de células Ki-64 positivas (indicando desarrollo de capilares) antes (C) y después (D) de 4 semanas de entrenamiento intenso (Jensen et al., 2004).
El efecto del ejercicio intenso (150% del VO2máx) fue positivo sobre la proliferación de células endoteliales en el músculo microdialisadotanto enla pierna entrenada como en la no entrenada (de 262 ± 60 al 573 ± 87 % del control de perfusión para la pierna no entrenada y de 303 ± 75 al 415 ± 108 % del control de perfusión para la pierna entrenada).
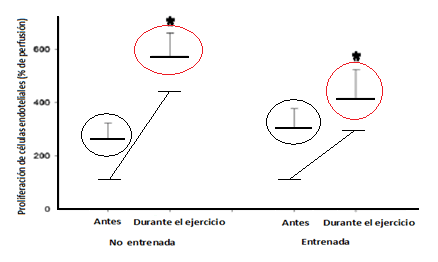 Figura 12. Proliferación de células endoteliales antes y durante el ejercicio fraccionado intenso (150% del VO2máx) para la pierna entrenada y la no entrenada. *P < 0.05 (modificado de Jensen et al., 2004).
Figura 12. Proliferación de células endoteliales antes y durante el ejercicio fraccionado intenso (150% del VO2máx) para la pierna entrenada y la no entrenada. *P < 0.05 (modificado de Jensen et al., 2004).
En el estudio se concluye que el ejercicio intermitente intenso induce a la formación de capilares y a la proliferación transitoria de células endoteliales en la fibra muscular, en magnitudes similares para las fibras tipo I y II.
Paralelamente al indicarse que la capilarización fibrilar no es una adaptación exclusiva de métodos de entrenamiento de resistencia tradicionales (como los continuos) y que los métodos de mayores intensidades podrían llegar a producir un mayor grado de capilarización fibrilar que los de menores, en este estudio se observa que las fibras tipo II también tienen capacidad de generar y aumentar capilares sanguíneos. Entonces se hipotetiza, debido a que los estudios como el presentado poseen protocolos diversos, que estas fibras poseerían una capacidad aeróbica tal como se viene observando. La incógnita a resolver es la relacionada con qué fibras sería preferible estimular en el marco de los deportes de equipo, teniendo en cuenta que ambas poseerían capacidades adaptativas aeróbicas.
¿CONTINUOS, INTERVALADOS, INTERMITENTES O SPRINTS INTERVALADOS-REPETIDOS?
Por lo presentado parecería que la evidencia científica indicaría que un aspecto relevante en el terreno de las adaptaciones fisiológicas, inclusive en el de las adaptaciones aeróbicas, es la intensidad del ejercicio. En esta dirección, Noakes & Sheehan (1991) señalan que los beneficios del entrenamiento dependen de la distancia cubierta a alta velocidad, esto determinaría las adaptaciones musculares maximizando el número de contracciones musculares potentes. Entonces, metodologías que permitan sostener una alta y máxima intensidad durante significativos periodos de tiempo podrían empezar a cobrar una radical importancia en el contexto de la preparación física deportiva actual. Esta ponderación es muy diferente a aquella que propone el alto volumen en desmedro de la intensidad. Este enfoque propone el alto volumen de alta y máxima intensidad.
Las metodologías más eficientes para conseguir el objetivo propuesto anteriormente en la actualidad podrían ser los intermitentes de distintos tipos (“Intermittent Training”) como bien lo evidencian distintos estudios (Astrand, I., et al., 1960a y 1960b; Casas, A., 2008) y distintos protocolos de sprints intervalados y repetidos. Por las características de estas 2 metodologías se puede deducir que ambas pueden combinar en mayor grado esta premisa de «alto volumen de alta y máxima intensidad» que los clásicos intervalados y continuos en especial.
Existen otras metodologías que pueden ser sumadas a la discusión como lo son los físico-técnicos, físico-tácticos o juegos reducidos, ya que también son metodologías que concuerdan con esta línea, permiten sumar y combinar esta idea de la alta intensidad con la mayor especificidad que el entrenamiento en la actualidad reclama. La misma es integradora de las metodologías anteriormente señaladas (intermitentes y sprints intervaldos y repetidos). En esta metodología se debería buscar que las intensidades sean altas, en estos casos se puede controlar a través de la FC (Frecuencia Cardíaca). En relación a esto MacMillan et al. (2005) analizaron las adaptaciones producidas por un protocolo de entrenamiento físico-técnico para el fútbol. Entrenaron durante 2 veces semanales, aparte de su entrenamiento semanal. El entrenamiento consistió en un circuito técnico en pista que mantenía una intensidad del 90-95% de la FCmáx durante los intervalos intensos (4 min) y del 70% de la FCmáx durante los períodos de recuperación activa (3 min). Se midió el VO2máx y los cm alcanzados en distintos saltos (SJ y CMJ).antes y después de un período de10 semanas de entrenamiento. En ambos casos se registraron aumentos significativos (P < 0.001 para el VO2máx y P < 0.05 para ambos saltos). En cuanto a las cargas y volúmenes a utilizar para este tipo de metodología, tomando en cuenta los resultados de este tipo de estudios, por el momento se podrían utilizar como parámetros generales las dosificaciones propuestas por los mismos. Sin embargo para proponer dosificaciones más concretas en este sentido aún hace falta más investigación.
Por último en esta línea también se puede sumar la metodología por circuitos (“Circuit Training”) ya que es abarcativa de las anteriores, donde las mismas pueden desarrollarse simultáneamente con la suma de otros estímulos de importancia para este tipo de disciplinas deportivas (los deportes de conjunto de dinámica intermitente), como los neuromusculares relacionados con la fuerza y la potencia con distintos tipos de trabajos de sobrecarga y pliométricos (Cometti, 2002; de la Reina Montero & Martínez de Haro, 2003). Una posibilidad es trabajar con 6 a 12 ejercicios de aproximadamente 30” deduración cada uno, con 2 a 3 vueltas por bloque o serie (de la Reina Montero & Martínez de Haro, 2003).
Estas metodologías también permitirían, junto a las adaptaciones aeróbicas, generar mayor cantidad de adaptaciones importantes especialmente para estos deportes, como lo pueden ser sobre las fibras rápidas tipo II (Cometti, G., 2002) que se ven especialmente estimuladas principalmente por el constante y sistemático freno y arranque, con aceleraciones y desaceleraciones, que en general estas metodologías proponen (Sassi, R., Rampinini, E., 2010). Como evidencian distintos estudios de electromiografía esta tipología de esfuerzo impone un especial estrés sobre los factores locales de la resistencia y las fibras tipo II (Borrani, F., et al., 2001; Greig, M. P., et al., 2006). Otros factores de importancia, en especial en este contexto, estimulados directamente por este tipo de metodologías, son los relacionados con la utilización específica del O2 mioglobínico (Astrand, I., et al., 1960b) entre otros como la eficiencia de los transportadores de glucosa (GLUT-4) que también se beneficia por el estímulo de elevada intensidad (Terada, S., et al., 2001). Estos factores están especialmente implicados en la competencia de estos deportes.
Los factores fisiológicos anteriormente señalados y demás relevantes en este marco son estimulados específica y directamente por este tipo de metodologías a diferencia de lo que sucede con el tradicional continuo y el clásico intervalado. Incluso el método por pasadas, que también es de características fraccionadas, propone un número muy inferior de micro pausas, frenos, arranques y de especificidad general que los intermitentes, sprints repetidos y físico-técnicos como los mencionados. Por estas razones no existen sólidos argumentos que justifiquen su utilización en remplazo, aunque sea temporalmente, de distintas propuestas que proponen mayor intensidad y especificidad en este contexto. El realizar este remplazo no sólo quitaría tiempo para generar adaptaciones útiles en mayor grado por causa del lapso más elevado de estímulo intenso y específico, sino que incluso podría generar desmejoras sobre algunos factores de importancia, como los fibrilares, debido a que en este aspecto esas metodologías acentuarían el estímulo sobre las fibras tipo I, por sus menores intensidades y frenos-arranques (Borrani et al., 2001), lo cual no sería muy conveniente en el marco de los deportes de equipo (Cometti, 2002). Se puede hipotetizar entonces que estas disminuciones a pesar de luego poder ser compensadas, probablemente afecten la posibilidad de que estos factores consigan llegar al nivel que pudieron haber alcanzado sin esta línea de base disminuida y con un mayor tiempo de estímulos más intensos, específicos y eficientes.
Un posible caso que pudiese justificar metodologías tradicionales (especialmente continuos) en este contexto,pudiese ser la necesidad de disminuir el tejido adiposo. Sin embargo, hay que recordar el papel que tendría la intensidad sobre el mecanismo de la lipólisis (desdoblamiento de los triglicéridos en ácidos grasos y glicerol). Es conocido el efecto que la adrenalina incrementada por la intensidad ejerce sobre el estímulo de la enzima encargada de catalizar el proceso de lipólisis (lipasa hormo sensible). A pesar de que la lipolisis no justifica la oxidación de todos los AG liberados, este efecto fisiológico, como otros como el mayor gasto energético que la alta intensidad genera incrementando el EPOC, podrían dar algunos principios justificativos para las disminuciones significativas del tejido adiposo producidas por protocolos cortos de sprints repetidos, según algunos estudios incluso en magnitudes superiores a los efectos producidos por continuos de mayores volúmenes (Trapp, 2006). El estudio de Trapp midió de forma directa el tejido adiposo (utilizando DEXA) antes y después de un protocolo de sprints repetidos y un protocolo de entrenamiento continuo.
Por estas razones la combinación de la alta y máxima intensidad con la posterior moderada/baja intensidad, integrando metodologías, pudiese resultar eficaz para estos casos, especialmente para facilitar la posterior combustión mitocondrial de los ácidos grasos que liberaría la alta intensidad, buscando evitar su re-esterificación. Esta argumentación podría resultar también aplicable para la justificación de los continuos variables (fartlek) en estos casos, que cabe destacar que son propuestas con características diferentes a los continuos extensivos y dignas de también ser tenidas en cuenta para el entrenamiento de la resistencia en este tipo de deportes por la característica variable de su esfuerzo, que puede responder en un nivel importante a la especificidad del esfuerzo de los mismos.
Según algunos autores los intermitentes también podrían ser una metodología a tener en cuenta desde este tema de la utilización de lípidos (Essén et al., 1977; Scott, 2002a y b; Kohan et al., 2008; Casas, 2008), pero esto no está aún muy claro y se encuentra en discusión. Son necesarios más estudios de investigación de mayor especificidad para poder abordar esta relación, entre los intermitentes y la utilización de lípidos, con mayores grados de precisión.
Por todo lo anteriormente señalado, sobre las adaptaciones aeróbicas sumadas a todos los estímulos adaptativos específicos mencionados (sobre aspectos anaeróbicos, energéticos, hormonales, enzimáticos y neuro-fibrilares), es que se podría considerar a este tipo de metodologías más intensas y específicas en relación alos deportes de equipo como el fútbol, rugby, hockey s/césped, etc. (intermitentes, sprints intervalados y repetidos, físico-técnicos, circuitos), como eficientes para el entrenamiento de la resistencia desde sus bases, especialmente en cuanto al aprovechamiento del tiempo, en el marco de los deportes de conjunto. Cabe destacar que la intención de este análisis no es desestimar por completo ninguna metodología, en especial debido a que la discusión sobre muchos de estos tópicos aún continúa abierta.
Cuando se habla de las bases generales no se debe olvidar que la mayoría de los autores que utilizan estos términos aclaran que esas bases deben ser en relación al deporte en cuestión, por lo tanto existirían bases generales para cada especificidad deportiva como directamente se entiende si se lee completamente estos fragmentos que aparecen en distintos escritos como el libro “La resistencia” de Navarro Valdivieso, F. (1999). Entonces, en el caso de las disciplinas del atletismo de carrera de fondo de alto volumen, el continuo extensivo podría ser la metodología principal para estimular las bases aeróbicas propias de dicha disciplina, mientras que en deportes de conjunto como el fútbol, los intermitentes aeróbicos “lineales”, ya que existe una gran variedad de intermitentes que se pueden utilizar, podrían ser los trabajos de base para este tipo de modalidades deportivas, para luego avanzar con una gran variedad de posibilidades de estas mismas propuestas y otras de mayores intensidades y especificidad como distintos tipos de sprints repetidos, de físico-técnicos, etc.
En los comienzos de los procesos de entrenamiento deberían ser adecuadamente ordenados y programados los protocolos y progresos de estas metodologías (intensidades, volúmenes, tiempo de trabajo, de pausa y la densidad de su relación) para no generar una desmedida sobrecarga inicial. Con este progreso metodológico el argumento sobre las posibles lesiones que este tipo de metodologías más intensas podrían producir en el inicio de un proceso de entrenamiento carecería de justificados sustentos. En relación a la información presentada y los análisis desarrollados se podría deducir que las metodologías de menores intensidades y más inespecíficas sólo podrían ser justificadas por un período muy breve de tiempo con el fin de no comenzar un proceso desde el primer día con estímulos fuertes y demaciado estresantes, predisponiendo negativamente a los deportistas y generando alguna posibilidad de lesión inicial por las desadaptaciones orgánicas globales con que generalmente los deportistas comienzan un proceso de entrenamiento físico. Sin embargo, como se ha expuesto, esto puede ser subsanado con el manejo de las cargas, que trabajos como los intermitentes permiten perfectamente (donde se pueden utilizar distintos volúmenes e intensidades que puedenpartir desde el 90-100 % de la VAM o menos inclusive si se lo desea). En el caso de las otras metodologías analizadas y propuestas como la de sprints intervaldos y sprints repetidos, también pueden ser controlados y manipulados sus volúmenes generales,tiempos de esfuerzos y pausas, densidad de trabajo, entre otras variables para garantizar un adecuado progreso metodológico.
CONCLUSIONES
Las metodologías fraccionadas de esfuerzos cortos que proponen mayores intensidades generarían tantas o mayores adaptaciones aeróbicas que las de menores intensidades y mayores volúmenes.
Las metodologías que imponen un mayor grado de especificidad para los deportes de conjunto de dinámica intermitente producirían mayores adaptaciones específicas, neuromusculares, anaeróbicas y aeróbicas, que otras metodologías menos específicas como lo pudiesen ser los tradicionales continuos e intervalados en el contexto de los deportes de equipo.
No existe evidencia científica sólida desde el punto de vista fisiológico que indique que las metodologías que proponen un mayor grado de intensidad y/o especificidad deberían ser precedidas por otras en este contexto deportivo.
Otras argumentaciones que en especial en este contexto tienden a justificar la utilización de períodos relativamente más extensos de metodologías de intensidades bajas y/o más inespecíficas de entrenamiento de la resistencia, no poseerían un consistente sustento científico en la actualidad.
REFERENCIAS
1. Andrienko, T., Kuznetsov, A.B., Kaambre, T., Usson, Y., Orosco, A., Appaix, F., Tiivel, T., Sikk, P., Vendelin, M., Margreiter R., Saks, V.A (2003). Metabolic consequences of functional complexes of mitochondria, myofibrils and sarcoplasmic reticulum in muscle cells. The Journal of Experimental Biology. 206, 2059-2072
2. Astrand I., Astrand P.O., Hedman, R (1960). Intermittent muscular work. Acta Physiological Scandinavian. 48, 443
3. Astrand I., Astrand P.O., Hedman, R (1960). Myohemoglobin as an Oxygen-stored in Man. Acta Physiological Scandinavian; 48, 454
4. Aziz, A.R., Mukherjee, S., Chia, M.Y.H., The, K.C (2008). Validity of the running repead sprint ability test among playing positions and level of compatitiveness in trained soccer players. Journal Sport Medicine. ISSN: 0172–4622
5. Baker, S.K., McCullagh K.J.A., Bonen A (1998). intensity-dependent and tissue-specific increases in lactate uptake and MCT-1 in heart and muscle. Journal Applied of Physiology; 84 (3), 987–994
6. Bangsbo, J., Iaia, F.M., Krustrup, P (2008). The Yo-Yo Intermittent Recovery Test: a useful tool for evaluation of physical performance in intermittent sports. Sports Medicine; 38 (1), 10112–1642
7. Barbero Álvarez, J. C.; Heredia Jiménez, J.M.; Méndez Villanueva, A (2005). Relación entre el YOYO test de recuperación intermitente y la capacidad de sprints repetidos en jugadores profesionales de deportes de equipo. Journal of Sports Sciencies; 23, 11–12
8. Barbero Álvarez, J. C.; Méndez Villanueva, A.; Bishop, D (2006). La capacidad para repetir esfuerzos máximos intermitentes: aspectos fisiológicos (I). Archivos de Medicina del Deporte; 114, 299–304
9. Barbero Álvarez, J. C.; Méndez Villanueva, A.; Bishop, D (2006). La capacidad para repetir esfuerzos máximos intermitentes: aspectos fisiológicos (II). Archivos de Medicina del Deporte; 115, 379–390
10. Barbours, R.L., Ribaudol, J., Chanoy, S.H.P (1984). Effect of creatine kinase activity on mitochondrial ADP/ATP transport. The Journal of Biological Chemestry; 259 (13): 8246–8251
11. Besseman, S.P., Geiger, P.J (1981). Ransport of energy in muscle: the phosphorylcreatine shuttle. Science; 211, 448–452
12. Bickham, D.C., Bentley, D.J., Le Rossignol,P.F., Cameron-Smit,D (2006). The effects of short-term sprint training on MCT expression in moderately endurance-trained runners. European Journal Applied of Physiology; 96 (6), 636–643
13. Borrani, F., Candau, R., Millet, G.Y., Perrey, S., Fuch-slocher, J., Rouillon, J.D (2001). Is the VO2 slow component dependent on progressive recruitment of fast-twitch fibers in trained runners?. Journal Applied Physiology; 90, 2212–2220
14. Brooks, G., Fahey, T., Baldwin K (2005). Exercise Physiology: Human bioenergetics and its applications. . (4ta Edición) McGraw Hillç
15. Burgomaster, K.A., Howarth, K.R., Phillips, S.M., Rakobowchuk, M., MacDonald, M.J., McGee, S.L., Gibala, M.J (2008). Similar metabolic adaptations during exercise after low volume sprint interval and traditional endurance training in humans. Journals of physiology; 586 (1), 151–160
16. Burgomaster, K.A., Cermak, N.M., Phillips, S.M., Benton C.R., Bonen, A., Gibala, M.J (2007). Divergent response of metabolite transport proteins inhuman skeletal muscle after sprint interval training and detraining. American Journal Physiology – Regulatory, Integrative, and Comparative Physiology;292, R1970–R1976
17. Burgomaster, K.A., Heigenhauser, G.J.F, Gibala, M.J (2006). Effect of short-term sprint interval training on human skeletal muscle carbohydrate metabolism during exercise and time-trial performance. Journal applied of physiology; 100 (6), 2041–2047
18. Burgomaster, K.A., Hughes, S.C., Heigenhauser, G.J.F., Bradwell, S.N., Gibala, M.J (2005). Six sessions of sprint interval training increases muscle oxidativepotential and cycle endurance capacity in humans. Journal of Applied Physiology;98, 1985–1990
19. Castagna, C., Abt, G., D’Ottavio S (2005). Competitive-level differences in yo-yo intermittent recovery and twelve minute run test performance in soccer referees. Journal of Strength and Conditioning Research; 19 (4), 805–809
20. Castagna, C.; Impellizzeri, F.M., Rampinini, E., D’Ottavio, S., Manzi, V (2008). The Yo–Yo intermittent recovery test in basketball players. Journal of Science and Medicine in Sport;.11, 202–208
21. Cometti, G (2002). La preparación física en el fútbol. Paidatribo
22. Coyle, E.F (2005). Very intense exercise-training is extremely potent and time efficient: a reminder. Journal of Applied Physiology; 98, 1983–1984
23. da Silva, J.F, Guglielmo, L.G.A, Bishop, D (2010). Relationship between different measures of aerobic fitness and repeated-sprint ability in elite soccer players. Journal of Strength and Conditioning Research; 24(x): 000–000
24. de la Reina, L., Martínez de Haro, M.V (2003). Manual de teoría y práctica del acondicionamiento físico. CV Ciencias del Deporte
25. Dubouchaud, H., Butterfield, G.E., Wolfel, E.E., Bergman, B.C., Brooks. G.A (2000). Endurance training, expression, and physiology of LDH, MCT1, and MCT4 in human skeletal muscle. American Journal of Physiology-Endocrinology and Metabolism; 278, E571–E579
26. Edge, J., Bisohop D., Hill-Haas S., Dawson B., Goodman C (2006). Comparison of muscle buffer capacity and repead-sprint ability of untrained, endurance-trained and team-sport atheletes. Journal Applied of Physiology; 96 (3), 225–234
27. Esper, R.C., Carrillo Córdova, J.R., Carrillo Córdova, L.D (2008). Modelo fisicoquímico del equilibrio ácido-base. Conceptos actuales (3ª de tres partes). Rev. Facultad de Medicina UNAM; 51 (5), 193–195
28. Essén, B., Hagenfeldt, L., Kaijser, L (1977). Utilization of blood-borne and intramuscular substrates during continuous and intermittent exercise in man. Journals of Physiology; 265, 489–506
29. Fernandez-Fernandez, J., Zimek, R., Wiewelhove, T., Ferrauti, A (2012). High-intensity interval training vs. Repeated-sprint training in tennis. Journal of Strength and Conditioning Research; 26 (1), 53–62
30. Ferrari Bravo, D., Impellizzeri, F.M., Rampinini, E., Castagna, C., Bishop, D., Wisloff, U (2008). Sprint vs. Interval Training in Football. Journal Sports Medicine; 29, 668–674
31. Finn, C (2001). Effects of High-Intensity Intermittent Training on Endurance Performance. Sportscience; 5 (1)
32. Freyssenet, D (2007). Energy sensing and regulation of gene expression in skeletal muscle. Journal applied of physiology; 102 (2), 529–540
33. Gaitanos, G.C., Williams C., Boobis L.H., Brooks S (1993). Human muscle metabolism during intermittent maximal exercise. Journal Applied Physiology; 75, 712–719
34. Gibala, M.G., Little, J.P., van Essen, M., Wilkin, G.P., Burgomaster, K.A., Safdar, A., Raha, S., Tarnopolsky, M.A (2006). Short-term sprint interval versus traditional endurance training: similar initial adaptations in human skeletal muscle and exercise performance. Journals of physiology; 575, 905–911
35. Gladden, L.B (2004). Lactate metabolism: a new paradigm for the third millennium. Journal of Physiology; 558 (1), 5–30
36. Green, H.J., Duhamel, T.A., Holloway, G.P., Moule J.W., Ranney, D.W., Tupling, A.R., Ouyang J. (2007). Rapid upregulation of GLUT-4 and MCT-4 expression during 16 h of heavy intermittent cycle exercise. American Journal Physiology-Regularoy, Integrative and Comparative Physiology; 294, R594–R600
37. Greig, M.P., MacNaughton, L.R., Lovell, R.J (2006). Physiological and mechanical response to soccer-specific intermittent activity and steady-state activity. Research in Sports Medicine; 14, 29–52
38. Harmer, A.R., McKenna, M.J., Sutton, J.R., Snow, R.J., Ruell, P.A., Booth, J., Thompson, M.W., Mackay, N.A., Stathis, C.G., Crameri, R.M.; Carey, M.F., Eager, D.M (2000). Skeletal muscle metabolic and ionic adaptations during intense exercise following sprint training in humans. Journal of Applied Physiology; 89, 1793–1803
39. Greig, M.P., MacNaughton, L.R., Lovell, R.J (2006). Physiological and mechanical response to soccer-specific intermittent activity and steady-state activity. Research in Sports Medicine; 14, 29–52
40. Huang, Y., Xiao, T., Liang, Y (2007). Progress in study of exercise, monocarboxylate transporter 1 and lactic shuttle. Life Science Journal; 4 (3), 57–73
41. Impellizzeri, F.M., Rampinini, E., Castagna, C., Bishop, C., Ferrari bravo, D., Tibaudi, A., Wisloff, U (2008). Validity of a repeated-Sprint test for football. Journal Sports Medicine. ISSN: 0172–4622
42. Impellizzeri, F.M., Rampinini, E., Castagna, C., Ferrari bravo, D., Bishop, C (2006). Capacità di ripeteregli sprint e suavalutazione. Rivista di Cultura Sportiva; 73, 31–34
43. Jensen, L., Bangsbo, J., Hellsten, Y (2004). Effect of high intensity training on capillarization and presence of angiogenic factors in human skeletal muscle. Journal of Applied Physiology; 557 (2), 571–582
44. Juel, C., Klarskov, C., Nielsen, J.J., Krustrup, P., Mohr M., Bangsbo, J (2003). Effect of high-intensity intermittent training on lactate and H+ release from human skeletal muscle. American Journal of Physiology – Endocrinology and Metabolism; 286, E245–E251
45. Kohan, A., Figueroa R., Dolce, P (2011). Comparación de variables fisiológicas y rendimiento alcanzado entre un patrón de ejercicio continuo vs un patrón de ejercicio intermitente en futbolistas profesionales [Internet], Futbol rendimiento. Disponible en: <http://futbolrendimiento.com.ar> [ Fecha de acceso 29 de Septiembre]
46. Krustrup, P., Bangsbo, J (2001). Physiological demands of top-class soccer refereeing in relation to physical capacity: effect of intense intermittent exercise training. Journal of Sports Sciences. 19, 881–891
47. Krustrup, P., Mohr, M., Amstrup, T., Rysgaard, T., Johansen, J., Steensberg, A., Pedersen, P.K., Bangsbo, J (2003). The Yo-Yo Intermittent Recovery Test: Physiological Response, Reliability, and Validity. Medicine & Science in Sports & Exercise; 35 (4), 697–705
48. Krustrup, P., Mohr, M., Ellingsgaard, H., Bangsbo, J (2005). Physical Demands during an Elite Female Soccer Game: Importance of Training Status. Medicine & Science in Sports & Exercise; 37 (7), 1242–1248
49. Krustrupo, P., Mohor, M., Nybo, L., Majgaard, J.J., Nielsen, J.J., Bangsbo, J (2006). The Yo-Yo IR2 Test: Physiological Response, Reliability, and Application to Elite Soccer. Medicine & Science in Sports & Exercise; 38 (9), 1666–1673ç
50. Little, J. P., Safdar, A., Wilkin, G. P., Tarnopolsky, M. A., Gibala M. J (2010). A practical model of low-volume high-intensity interval training induces mitochondrial biogenesis in human skeletal muscle: potential mechanisms. Journal of Physiology. 588 (6), 1011–1022
51. MacDougall, D. J., Hicks, A. L., MacDonald, J. R., McKelvie, R. S., Green, H. J., Smith, K. M (1998). Muscle performance and enzymatic adaptations to sprint interval training. Journal of Applied Physiology. 84 (6): 2138–2142
52. MacDougall, D. J., Hicks, A. L., MacDonald, J. R., McKelvie, R. S., Green, H. J., Smith, K. M (2004). Rendimiento muscular y adaptaciones enzimáticas al entrenamiento Intervalado de sprint. PubliCE <http://www.sobreentrenamiento.com>. Pid: 248
53. McMillan, K., Helgerud, J., Macdonald, R., Hoff, J (2005). Physiological adaptations to soccer specific endurance training in professional youth soccer players. Journal Sports Medicine; 39, 273–277
54. Martin, D., Carl, K., Lehnertz, K (2001). Manual de metodología del entrenamiento deportivo. Paidatribo
55. Mendoza Medellín, A (2008). El origen de la acidez en la glucólisis. REB; 27 (4), 111–118
56. Messonnier, L., Kristensen, M., Juel, C., Denis C (2007). Importance of pH regulation and lactate/H+ transport capacity for work production during supramaximal exercise in humans. Journal Applied of Physiology; 102, 1936–1944
57. Navarro Valdivieso, F (1999). La Resistencia. Gymnos
58. Noakes, T.; Sheehan, G (1991). Lore of running. Champaign (IL): Leisure Press, cop
59. Pilegaard, H., Domino, K., Noland, T., Juel, C., Hellsten, Y., Halestrap, A.P., Bangsbo J (1999). Effect of high-intensity exercise training on lactate/H1 transport capacity in human skeletal muscle. American Journal the Physiology-Endocrinology and Metabolism; 276 (39), E255–E261
60. Prakash, E. S., Robergs, R.A., Miller, B.F., Gladden, L.B., Jones, N., Stringer, W. W., Wasserman, K., Moll, W., Gros, G., Rowlands, D.S., Sahlin, K., Beneke, R (2008). Comments on Point: Counterpoint “Lactic acid is/is not the only physicochemical contributor to the acidosis of exercise. JournalApplied of Physiology; 105, 363–367
61. Robergs, R (2001). Exercise-Induced Metabolic Acidosis: Where do the protons come from?. Sportsciencie; 5 (2)
62. Robergs, R., Farzenah, G., Daryl, P (2004). Biochemistry of exercise-induced metabolic acidosis. American Journals of Physiology; 287, R502–R516,
63. Rodas, G., Ventura, J.L, Cadefau, J.A., Cusso, R., Parra, J (2000). A short training programme for the rapid improvement of both aerobic and anaerobic metabolism. European Journal of Applied Physiology; 82, 480–486
64. Rodas, G., Ventura, J.L., Cadefau, J.A., Cusso, R., Parra, J (2000). A short training programme for the rapid improvement of both aerobic and anaerobic metabolism. European journal of applied physiology; 82 (5-6), 480–486
65. Sassi, R., Rampinini, E (2011). La fase dimenticata: quelladelladecelerazione [Internet], Alleniamo. Disponible en <http://www.alleniamo.com> [Fecha de acceso 29 de Septiembre]
66. Savabi, F., Carpenter, C.L., Mohan, C., Besseman S.L (1998). The polysome as a terminal for the creatine phosphate energy shuttle. Biochemical Medicine and Metabolic Biology; 40 (3), 291–8
67. Schnyder, T., Gross, H., Winkler., H., Eppenberger, H.M., Walliman T (1991). Structure of the mitochondrial creatine kinase octamer: high-resolution shadowing and image averaging of single molecules and formation of linear filaments under specific staining conditions. The Journal of Cell Biology; 112 (1), 95–101
68. Scott, R (2002). Intermittent training aerobic training (Part 1). American Fitness; 63–64
69. Scott, R (2002). Intermittent training aerobic training (Part 2). American Fitness; 67–68
70. Spencer, M.,Bishop, D.,Dawson, B.,Goodman, C (2005). Physiological and Metabolic Responses of Repeated-Sprint Activities Specific to Field-Based Team Sports. Sports Medicine; 35 (12), 1025–1044
71. Stepto, N.K., Hawley, J.A., Dennis, S.C., Hopkins, W.G (1999). Effects of different interval-training programs on cycling time-trial performance. and Science in Sports and Exercise; 31, 735–741
72. Tabata, I., Nishimura, K., Kouzaki, M., Yuusuke, H., Ogita, F., Miyachi, M., Yamamoto, K (1996). Effects of moderate-intensity endurance and hiintensity intermittent training on anaerobic capacity and [spacing dot above] VO2máx. & Science in Sports & Exercise; 28 (10), 1327–1330
73. Terada, S., Yokozeki, T., Kawanaka, K., Ogawa, K., Higuchi, M., Ezaki, O., Tabata, I (2001). Effects of high-intensity swimming trainingon GLUT-4 and glucose transport activity in rat skeletalmuscle. Journal of Applied Physiology;90, 2019–2024
74. Thomas, C., Bishop, D., Moore-Morris, T., Mercier, J (2007). Effects of high-intensity training on MCT1, MCT4, and NBC expressions in rat skeletal muscles: influence of chronic metabolic alkalosis. American Journal of Physiology – Endocrinology and Metabolism; 293 (4), E916–22
75. Trapp, E.G (2006). Effect of high intensity exercise on fat loss in young overweight women. The University of New South Wales, School of Medical Sciences Health and Exercise Science
76. Vendelin, M., Lemba, M., Saks, V.A (2004). Analysis of functional coupling: mitochondrial creatine kinase and adenine nucleotide translocase. Biophysical Journal; 87, 696–713
77. Wallimann, T., Wyss, M.S., Brdiczka, D., Klaas, N.T., Eppenberger, H.M (1992). Intracellular compartmentation, structure and function of creatinekinaseisoenzymes in tissues with high and fluctuating energy demands: the phosphocreatine circuit' for cellular energy homeostasis. Biochemical Journal; 281, 21–40
Efectos del entrenamiento combinado realizado dos o cuatro veces por semana sobre la presión arterial de 24 horas, hemoglobina glucosilada y otros resultados relacionados con la salud en personas mayores con hipertensión: Justificación y protocolo del estudio de un ensayo clínico aleatorio
Efectos del entrenamiento combinado en hipertensión y salud en adultos mayores El entrenamiento...
Efectos del ejercicio aeróbico y del ejercicio de resistencia sobre los índices físicos y los factores de riesgo cardiovascular en niños en edad escolar con sobrepeso y obesidad: una revisión sistemática y un metaanálisis
Ejercicio Aeróbico y de Resistencia: Soluciones para la Obesidad Infantil La obesidad infantil es...
Problemas de bienestar: Su influencia en la RPE y el disfrute en el entrenamiento de waterpolo máster y la brecha jugador–entrenador
El estudio reciente sobre el bienestar y sus efectos en la carga de entrenamiento y disfrute en el...
Efectos del ejercicio en la composición corporal de pacientes con cáncer de próstata que reciben terapia de privación androgénica: una revisión sistemática actualizada y un metaanálisis
Efectos del ejercicio en la composición corporal de pacientes con cáncer de próstata bajo terapia...
Aumento inducido por el ejercicio en el factor neurotrófico derivado del cerebro (BDNF) en personas con esclerosis múltiple: Una revisión sistemática y metaanálisis de ensayos de intervención de ejercicio
El papel del Factor Neurotrófico Derivado del Cerebro (BDNF) en la esclerosis múltiple (EM) ha...