
Exercise & Sport Nutrition Laboratory, Department of Human Movement Sciences & Education, The University of Memphis, Memphis, TN.
Resumen
ANTECEDENTES
Los aminoácidos son los bloques con los cuales se construyen las proteínas corporales, como tales son esenciales para la síntesis de proteínas estructurales, enzimas y algunas hormonas y neurotransmisores. Los aminoácidos están también involucrados en numerosas vías metabólicas que afectan el metabolismo durante el ejercicio. Consecuentemente, se ha sugerido que los atletas involucrados en entrenamientos intensos requieren proteínas adicionales en su dieta o que deberían suplementar sus dietas con aminoácidos específicos. Aquí he revisado los principios y la evidencia de los potenciales efectos ergogénicos de la suplementación a corto plazo con proteínas y aminoácidos y la evidencia de los potenciales efectos anabólicos de la suplementación a largo plazo cuando se la combina con el entrenamiento. He tratado primero las proteínas y luego los aminoácidos bajo los siguientes encabezados: los aminoácidos potencialmente anabólicos; los aminoácidos de cadena ramificada que tienen diferentes roles en el metabolismo y su potencial efecto sobre el rendimiento; la glutamina, que pertenece a una clase en si misma, por sus efectos sobre el sistema inmune; la creatina, un aminoácido que no es uno de los bloques con los cuales se construyen las proteínas, pero esta implicado en la producción muscular de energía a corto plazo; y el hidroximetilbutirato (HMB), un metabolito de la leucina potencialmente anabólico.
LITERATURA
Esta revisión es mas una actualización que un recuento exhaustivo de todos los trabajos publicados sobre este tema. He citado dos libros, 60 artículos de investigación, 10 resúmenes publicados, y 18 artículos de revisión/capítulos de libros de mi propia base de datos de referencias. En mi base de datos hay un adicional de 97 artículos de investigación, 78 resúmenes, y 38 artículos de revisión o capítulos de libros sobre este tema. Estas referencias adicionales están revisadas en otros artículos (Kreider, 1999, Kreider 1998; Williams, et al, 1999).
HALLAZGOS
Proteínas
Una considerable cantidad de investigaciones han evaluado las necesidades proteicas en las dietas de los atletas. Aunque existe cierto debate, la mayoría de los estudios indican que con el propósito de mantener el balance proteico durante el ejercicio de sobrecarga y/o ejercicios de resistencia, los atletas deberían ingerir aproximadamente 1.3 a 1.8 gramos de proteínas por kg de masa corporal por día (Butterfield, 1991; Lemon, 1998; Kreider et al., 1993; Kreider, 1999). Los atletas que entrenan en la altura podrían necesitar tanto como 2.2 gramos de proteínas por día con el propósito de mantener el equilibrio proteico (Butterfield, 1991). Esta ingesta proteica es casi 1.5 a 2 veces la recomendación dietética diaria (RDA) para los adultos normales. La mayoría de las veces una dieta iso – calórica puede proporcionar la cantidad requerida de proteínas, pero los atletas que mantengan dietas hipo – calóricas, no ingieren suficientes proteínas de calidad en su dieta, y/o entrenan en la altura, pueden ser susceptibles de malnutrición proteica (Kreider, 1999). En teoría, este estado puede enlentecer el crecimiento de los tejidos y/o la recuperación del entrenamiento. Por otro lado, ingerir mas proteínas de las necesarias para mantener el equilibrio proteico durante el entrenamiento (e.g., > 1.8 g/kg/día) no promoverá mayores ganancias en la fuerza o en la masa libre de grasa (Lemon et al., 1992; Tarnopolsky et al., 1992). Estos hallazgos indican que los atletas no necesitan suplementar sus dietas normales con proteínas, los atletas deberían ingerir proteínas de calidad suficiente para mantener el equilibrio proteico.
Mas recientemente, ha habido un interés en determinar los efectos las respuestas hormonales al ejercicio de las ingesta pre y post ejercicio con carbohidratos y proteínas (Cade et al., 1992; Chandler et al., 1994; Roy and Tarnopolsky, 1998; Tarnopolsky et al., 1997; Zawadzki et al., 1992). Se ha reportado que la ingesta de carbohidratos y proteínas incrementa los niveles de insulina y/o de la hormona de crecimiento a un mayor grado que la ingestión de carbohidratos por si sola (Chandler et al, 1994, Zawadzki et al, 1992). Consecuentemente, la ingesta de proteínas y carbohidratos previo al ejercicio puede servir como una estrategia nutricional anti catabólica (Carli et al, 1992). Además, la ingesta de carbohidratos y proteínas luego del ejercicio puede promover un perfil hormonal mas anabólico, la síntesis de glucógeno, y/o acelerar la recuperación luego del ejercicio intenso (Roy y Tarnopolsky, 1998, Roy et al, 1997). Con el tiempo estas alteraciones le darán al atleta una mayor tolerancia al entrenamiento y/o promoverá mayores adaptaciones al mismo, sin embargo la evidencia aun no es clara.
AMINOACIDOS ANABOLICOS
Uno de los beneficios comúnmente supuestos de la suplementación con aminoácidos es que ciertos aminoácidos (e.g., arginina, histidina, lisina, metionina, ornitina y fenilalanina) pueden estimular la liberación de la hormona de crecimiento, de la insulina y/o de los glucocorticoides, y de esta manera promover los procesos anabólicos (Kreider, 1993). Existe aluna evidencia clínica de que la suplementación con aminoácidos puede estimular la liberación de factores crecimiento y/o la liberación de la hormona del crecimiento (Carlson, et al., 1989;; Garlick and Grant, 1988; Iwasaki et al., 1987; Merimee et al., 1969). Por ejemplo, la infusión intravenosa de arginina y ornitina ha sido utilizada clínicamente para estimular la liberación de la hormona del crecimiento (Carlson et al., 1989; Iwasaki et al., 1987). Asimismo, estudios clínicos preliminares indican que las proteínas (20 a 60 gr); la arginina y la lisina (1.2 gr) y la ornitina (70 mg/kg) incrementan las concentraciones de hormona de crecimiento y de somatomedinas en la sangre (Bucci et al., 1990; Jackson et al., 1968; Isidori et al., 1981). Sin embargo, otros investigadores no han replicado estos hallazgos, particularmente en individuos sanos (Lemon, 1991). También hay poca evidencia de que la suplementación con estos aminoácidos durante el entrenamiento afecte significativamente la composición corporal, la fuerza y/o la hipertrofia muscular (Kreider, 1999). Consecuentemente, los efectos de la suplementación con aminoácidos sobre la liberación de la hormona del crecimiento y las adaptaciones al entrenamiento son aun inciertos.
AMINOACIDOS DE CADENA RAMIFICADA
Los investigadores han realizado un esfuerzo considerable evaluando los efectos de la suplementación con aminoácidos de cadena ramificada (AACC: lucina, isoleucina y valina) sobre las respuestas psicológicas y fisiológicas al ejercicio (Blomstrand et al., 1991; Kreider, 1998; Wagenmakers, 1998). Existen dos hipótesis principales con respecto al valor ergogénico de la suplementación con estos aminoácidos.
Primero, se ha reportado que la suplementación con AACR reduce la degradación proteica inducida por el ejercicio y/o la liberación de enzimas musculares (un indicador del daño muscular) posiblemente promoviendo un perfil hormonal anti catabólico (Carli et al., 1992; Coombes and McNaughton, 1995). Teóricamente, la suplementación con AACR durante el entrenamiento intenso puede ayudar a minimizar la degradación proteica y por lo tanto conducir a una mayor ganancia de masa libre de grasa. Aunque varios estudios respaldan esta hipótesis, se necesitan investigaciones adicionales para determinar los efectos a largo plazo de la suplementación con AACR durante el entrenamiento sobre los marcadores del catabolismo, la composición corporal y la fuerza (Kreider, 1998).
Segundo, se ha teorizado que la disponibilidad de AACR durante el ejercicio contribuye a la fatiga central (Newsholme et al, 1991). Durante ejercicios de resistencia, los AACR son absorbidos mas por los músculos que por el hígado con el propósito de contribuir al metabolismo oxidativo. La fuente de AACR para el metabolismo oxidativo muscular durante el ejercicio es la reserva plasmática de AACR, la cual es repletada a través del catabolismo corporal total de proteínas durante el ejercicio de resistencia (Davis, 1995; Kreider, 1998; Newsholme et al., 1991). Sin embargo, la oxidación de AACR en el músculo durante el ejercicio prolongado puede exceder la capacidad catabólica para incrementar la disponibilidad de AACR, por lo cual la concentración plasmática de los mismo puede disminuir durante el ejercicio de resistencia prolongado (Blomstrand et al., 1988; Blomstrand et al., 1991). La disminución en los AACR durante el ejercicio de resistencia puede resultar en un incremento del índice triptofano libre / AACR. El triptofano libre y los AACR compiten para entrar en el cerebro a través de un transportador de aminoácidos (Newsholme et al, 1991). De esta manera, la disminución en los AACR en la sangre facilita la entrada de triptofano al cerebro. Además, la mayoría del triptofano está unido a la albúmina, y la proporción de triptofano unido a la albúmina está influenciada por la disponibilidad de ácidos grasos de cadena larga (Davis et al., 1992; Newsholme et al., 1991). Durante el ejercicio de resistencia la concentración de ácidos grasos libres se incrementa, por lo cual la cantidad de triptofano unido a la albúmina cae, incrementando la concentración de triptofano libre en la sangre (Davis, 1995). Conjuntamente, la disminución en los AACR plasmáticos y el incremento del triptofano libre durante el ejercicio prolongado de resistencia altera el índice triptofano libre / AACR por lo cual se incrementa la entrada de triptofano al cerebro (Newsholme et al, 1991). La incrementada concentración de triptofano en el cerebro promueve la formación del neurotransmisor 5-hidroxitriptamina (%-HT). En estudios con animales y con humanos se ha mostrado que la 5-HT induce el sueño, la depresión de la excitabilidad de las motoneuronas, influencia las funciones autonómica y endocrina, y suprime el apetito.
El desequilibrio en el índice triptofano libre / AACR ha sido implicado como una posible causa de fatiga aguda fisiológica y psicológica (fatiga central). También se ha hipotetizado que el aumento crónico en la concentración de 5-HT, la cual puede ocurrir en atletas que mantienen un elevado volumen de entrenamiento, algunos de los signos y síntomas reportados del síndrome de sobreentrenamiento: hipotensión postural, anemia, amenorrea, inmunosupresión, supresión del apetito, perdida de peso, depresión y disminución del rendimiento (Newsholme et al., 1991; Gastmann and Lehmann, 1998; Kreider, 1998).
Recientemente se han conducido un cierto número de estudios para evaluar si la suplementación con carbohidratos o con AACR afecta la fatiga central durante el ejercicio y/o los signos y síntomas del sobreentrenamiento. El análisis de esta literatura indica que la suplementación con carbohidratos y/o AACR puede afectar el índice triptofano libre / AACR. Por ejemplo, se ha reportado que la administración de carbohidratos durante el ejercicio atenúa la liberación de FFA y minimiza el incremento del índice triptofano libre / AACR (Davis et al, 1993). Además, se ha reportado que la suplementación con AACR incrementa la concentración plasmática de los mismos y minimiza y/o previene el incremento en el índice triptofano libre / AACR (Blomstrand et al, 1991). También hay estudios que indican que la administración de AACR con o sin carbohidratos previo y durante el ejercicio puede afectar las respuestas fisiológicas y psicológicas al mismo (Coombes and McNaughton, 1995; Hefler et al., 1993; Kreider et al., 1992; Kreider and Jackson, 1994).
Sin embargo, aun no esta claro el efecto de estas alteraciones en el índice inducidas nutricionalmente triptofano libre / AACR sobre el rendimiento físico. La mayoría de los estudios indican que la suplementación con AACR no mejora el rendimiento en una serie única de ejercicio de resistencia, aunque sin dudas a estos estudios les faltó habilidad para delimitar pequeñas pero útiles mejoras en el rendimiento (Davis, 1995; Gastmann and Lehmann, 1998; Kreider, 1998). Es necesario realizar mas investigaciones para determinar el efecto a largo plazo de la suplementación con AACR sobre las adaptaciones al entrenamiento y los signos y síntomas del sobreentrenamiento (Kreider, 1998).
Glutamina
Rennie y cols han sugerido a la suplementación con glutamina como una estrategia para promover el crecimiento muscular (Rennie et al., 1994; Rennie, 1996). Estos autores se basaron en la sugerencias realizadas en estudios con animales y con humanos sobre los efectos de la glutamina sobre la síntesis proteica, el volumen celular, y la síntesis de glucógeno (Rennie et al., 1994; Varnier et al., 1995; Rennie, 1996; Low et al., 1996). La glutamina es también un combustible importante para las células blancas, por lo cual la reducción en la concentración sanguínea de glutamina luego del ejercicio intenso puede contribuir a la supresión inmune en atletas sobreentrenados (Parry-Billings et al., 1990a; Parry-Billings et al., 1990b; Parry-Billings et al., 1992; Kargotich et al., 1996; Newsholme and Calder, 1997).
Estudios preliminares indican que la suplementación con aminoácidos de cadena ramificada (4 a 16 gr) y/o glutamina (4 a 12 gr) puede prevenir la reducción o incluso incrementar la concentración de glutamina durante el ejercicio (Kreider, 1998). En teoría estos cambios en la concentración de glutamina podrían tener efectos beneficiosos sobre la síntesis proteica y la función inmune. Sin embargo, en los pocos estudio donde se halló un incremento en la disponibilidad de glutamina, hubo poco o ningún efecto sobre el rendimiento o el estátus inmune (Rohde et al., 1998; Nieman and Pedersen, 1999). También es poco claro si la suplementación a largo plazo con glutamina afecta la síntesis proteica, la composición corporal o la incidencia de infecciones en el tracto respiratorio superior durante el entrenamiento.
Creatina
La creatina es una aminoácido natural derivado de los aminoácidos glicina,
arginina y metionina (Balsom et al., 1994; Williams et al., 1999). La mayor
parte de la creatina esta almacenada en los músculos esqueléticos,
principalmente como fosfocreatina; el resto se halla en el corazón, el cerebro,
y en los testículos (Balsom et al., 1994; Kreider, 1998). El requerimiento
diario de creatina es de aproximadamente 2 a 3 gr; la mitad se obtiene de la
dieta, principalmente de la carne y del pescado, mientras que el resto es
sintetizado (Williams et al, 1999). Se ha propuesto a la suplementación con
creatina como una forma de “cargar” al músculo con creatina y con fosfocreatina
(PCr). En teoría, una reserva incrementada de creatina y fosfocreatina mejoraría
la habilidad para producir energía durante el ejercicio de alta intensidad, así
como también mejoraría la velocidad de recuperación luego del ejercicio de alta
intensidad.
Se han llevado a cabo un número de estudios para determinar los efectos de la
suplementación con creatina sobre las concentraciones musculares y sobre el
rendimiento. Se ha reportado que la suplementación con creatina (20 gr por día o
0.3 gr por kg de masa corporal por día durante 4 a 7 días) incrementa el
contenido intramuscular de creatina y de fosfocreatina en un 10 a un 30% (Casey
et al., 1996; Febbraio et al, 1995; Green et al., 1996a; Green et al., 1996b;
Greenhaff et al., 1993a; Hultman et al., 1996; Smith et al., 1998b, Vandenberghe
et al., 1997). Existe también evidencia de que la suplementación con creatina
mejora la tasa de resíntesis de PCr luego del ejercicio intenso (Greenhaff et
al., 1993b; Greenhaff et al., 1994a; Greenhaff et al., 1994b). La mayoría de los
estudios indican que la suplementación con creatina a corto plazo incrementa la
masa corporal total (Hultman et al., 1996; Williams et al., 1999), el
rendimiento durante series múltiples de contracciones musculares máximas (Greenhaff
et al., 1993a; Volek et al., 1997), y la capacidad para realizar un sprint único
y/o sprints repetidos (Birch et al., 1994; Grindstaff et al., 1997; Prevost et
al., 1997). Además, se ha reportado que la suplementación con creatina a largo
plazo promueve mayores ganancias en la fuerza (Earnest et al., 1995; Peeters et
al., 1999; Stone et al., 1999; Vandenberghe et al., 1997), en la masa libre de
grasa (Kreider et al., 1998; Stone et al., 1999; Stout et al., 1999;
Vandenberghe et al., 1997) y en el rendimiento en el sprint (Kreider et al.,
1998; Peyreburne et al., 1998; Stout et al., 1999). Sin embargo, debe señalarse
que no todos los estudios reportan beneficios ergogénicos (Burke et al., 1996;
Redondo et al., 1996; Terrillion et al., 1997), y se ha reportado que la cafeína
neutraliza el potencial valor ergogénico de la suplementación con creatina (Vanakoski
et al., 1998; Vandenburghe et al., 1996). Aunque es necesaria mayor
investigación, la suplementación con creatina parece ser una estrategia
nutricional segura y efectiva para mejorar el rendimiento en el ejercicio de
alta intensidad y para mejorar las adaptaciones al entrenamiento (Williams et
al., 1999).
Hidroximetilbutirato (HMB)
El metabolito de la leucina, el hidroximetilbutirato (mas exactamente la sal de calcio del ácido β hidroxi β metilbutirico) se ha vuelto recientemente un suplemento dietario popular que supuestamente promueve ganancias en la masa libre de grasa y en la fuerza durante el entrenamiento de sobrecarga (Kreider, 1999). La razón de esto es que la leucina y su metabolito el α-cetoisocaproato (KIC) parece inhibir la degradación proteica (Nair et al., 1992; Nissen et al., 1996) y su efecto anti proteolítico puede estar mediado por el HMB. Estudios con animales indican que aproximadamente el 5% de la leucina oxidada es convertida en HMB a través del KIC (Nissen et al., 1994; Van Koevering et al., 1994). La adición de HMB a una comida a base de colostrum de leche y de soja (Nissen et al, 1994), tendió a mejorar la calidad esquelética de potrillos (Van Koevering et al., 1994) y a disminuir los marcadores del catabolismo durante el entrenamiento en caballos (Miller et al., 1997). La suplementación con leucina y/o HMB puede de esta manera inhibir la degradación proteica durante los períodos asociados al incremento de la proteolisis, tal como el entrenamiento de sobrecarga.
Aunque mucha de la literatura disponible sobre la suplementación con HMB en humanos es preliminar, varios artículos y resúmenes recientemente publicados respaldan esta hipótesis. La infusión con leucina parece disminuir la degradación proteica en humanos (Nair et al, 1992). Se ha reportado que la suplementación con HMB durante 3 a 8 semanas de entrenamiento promueve ganancias significativamente mayores en la masa libre de grasa y en la fuerza en hombres y mujeres desentrenados que iniciaron un programa de entrenamiento de sobrecarga (Nissen et al., 1996; Nissen et al., 1997; Vukovich et al., 1997). En algunas instancia estas ganancias estuvieron asociada con signos significativos de menor daño muscular (flujo de enzimas musculares y excreción urinaria de 3-metilhistidina) (Nissen et al, 1996). Aunque estos hallazgos sugieren que la suplementación con HMB durante el entrenamiento puede mejorar las adaptaciones al mismo en individuos desentrenados que inician el entrenamiento, es menos claro si la suplementación con HMB reduce los marcadores del catabolismo o promueve mayores ganancias de masa libre de grasa y de fuerza durante el entrenamiento de sobrecarga en atletas bien entrenados. En efecto, existen varios reportes de efectos no significativos de la suplementación con HMB (3 a 6 gr por día) en atletas bien entrenados (Almada et al., 1997; Kreider et al., 1997; Kreider et al., 1999). Es necesaria mayor investigación (Kreider, 199).
CONCLUSIONES
La suplementación dietaria con proteínas mas allá de la necesaria para mantener el balance nitrogenado no proporciona un beneficio ergogénico adicional.
La ingesta de carbohidratos/proteínas previa al ejercicio puede reducir el catabolismo mientras que la ingesta de carbohidratos/proteínas luego del ejercicio puede promover la resíntesis de glucógeno, un ambiente hormonal mas anabólico, y la recuperación. El alcance en que estas estrategias afectan las adaptaciones al entrenamiento es desconocido.
Existe alguna evidencia extraída de ciertas poblaciones clínicas de que ciertos aminoácidos (e.g., arginina, histidina, lisina, metionina, ornitina y fenilalanina) pueden estimular la liberación de la hormona del crecimiento, la insulina y/o de los glucocorticoides y de esta manera promover los procesos anabólicos, sin embargo, hay poca evidencia de que la suplementación con estos aminoácidos proporcione algún efecto ergogénico para los atletas.
Se ha hipotetizado que los aminoácidos de cadena ramificada y la glutamina afectan la fatiga central y la supresión inmune inducida por el ejercicio, pero su valor ergogénico durante el ejercicio prolongado hasta el momento es confuso.
La mayoría de los estudios indican que la suplementación con creatina puede ser una manera efectiva y segura de mejorar el rendimiento en el ejercicio intermitente de alta intensidad así como también de mejorar las adaptaciones al entrenamiento. de los nutrientes evaluados, la creatina parece tener el mayor potencial ergogénico par los atletas involucrados en este tipo de entrenamiento.
Se ha reportado que la suplementación con hidroximetilbutirato reduce el catabolismo y promueve una mayor ganancia de fuerza y de masa libre de grasa en individuo desentrenados que inician el entrenamiento. Hay limitados datos disponibles sobre la suplementación con HMB sobre las adaptaciones al entrenamiento en atletas.
INVESTIGACIONES ADICIONALES
A lo largo de las últimas décadas los investigadores han hallado que los aminoácidos desempeñan múltiples roles en el metabolismo. Por esta razón, los investigadores y los atletas están interesados en los efectos de la suplementación con aminoácidos sobre el metabolismo durante el ejercicio, el rendimiento en el ejercicio y las adaptaciones al entrenamiento. Aunque se han realizado avances significativos, todavía queda mucho que aprender acerca de estos efectos. Los investigadores deberían también evaluar la seguridad a largo plazo de la suplementación con aminoácidos, así como también el potencial valor médico en el tratamiento de varias enfermedades.
REFERENCIAS
1. Cade JR, Reese RH, Privette RM et al (1992). Dietary intervention and training in swimmers. European Journal of Applied Physiology 63, 210-15
2. Butterfield G (1992). Amino acids and high protein diets. Perspectives in exercise science and sports medicine, Vol. 4; Ergogenics, enhancement of performance in exercise and sport (pages 87-122)
3. Carli G, Bonifazi M, Lodi L et al (1992). Changes in exercise-induced hormone response to branched chain amino acid administration. European Journal of Applied Physiology 64, 272-7
4. Chandler RM, Byrne HK, Patterson JG et al (1940). Dietary supplements affect the anabolic hormones after weight-training exercise. Journal of Applied Physiology 76, 839-45
5. Kreider RB (1999). Dietary supplements and the promotion of muscle growth with resistance training. Sports Medicine 27, 97-110
6. Kreider RB, Miriel V, Bertun E (1993). Aminoacid supplementation and exercise performance: proposed ergogenic value. Sports Medicine 16, 190-209
7. Lemon PW, Tarnopolsky MA, MacDougall JD et al (1992). Protein requirements and muscle mass/strength changes during intensive training in novice bodybuilders. Journal of Applied Physiology 73, 767-75
8. Lemon PWR (1998). Effects of exercise on dietary protein requirements. International Journal of Sport Nutrition 8, 426-47
9. Roy BD, Tarnopolsky MA (1998). Influence of differing macronutrient intakes on muscle glycogen resynthesis after resistance exercise. Journal of Applied Physiology 84, 890-96
10. Roy BD, Tarnopolsky MA, MacDougall JD et al (1997). Effect of glucose supplementation timing on protein metabolism after resistance training. Journal of Applied Physiology 82, 1882-88
11. Tarnopolsky MA, Atkinson SA ,MacDougall JD et al (1992). Evaluation of protein requirements for trained strength athletes. Journal of Applied Physiology 73, 1986-95
12. Tarnopolsky MA, Bosman M, acdonald JR et al (1997). Postexercise protein-carbohydrate and carbohydrate supplements increase muscle glycogen in men and women. Journal of Applied Physiology 83, 1877-83
13. Zawadzki KM, Yaspelkis BB, Ivy JL (1992). Carbohydrate-protein complex increases the rate of muscle glycogen storage after exercise. Journal of Applied Physiology 72, 1854-9
14. Bucci L, Hickson JF, Pivarnik JM et al (1990). Ornithine ingestion and growth hormone release in bodybuilders. Nutrition Research 10, 239-45
15. Carlson HE, Miglietta JT, Roginsky MS et al (1989). Stimulation of pituitary hormone secretion by neurotransmitter aminoacids in humans. Metabolism 28, 1179-82
16. Garlick PJ, Grant I (1988). Amino acid infusion increases the sensitivity of muscle protein synthesis in vivo to insulin. Biochemistry Journal 254, 579-84
17. Isidori A, Lo Monaco A, Cappa M (1981). A study of growth hormone release in man after oral administration of aminoacids. Current Medical Research Opinion 74, 75-81
18. Iwasaki K, Mano K, Ishihara M et al (1987). Effects of ornithine or arginine administration on serum amino acid levels. Biochemistry International 14, 971-6
19. Jackson D, Grant DB, Clayton B (1968). A simple oral test of growth hormone secretion in children. Lancet 2, 373-5
20. Kreider RB (1999). Dietary supplements and the promotion of muscle growth with resistance training. Sports Medicine 27, 97-110
21. Kreider RB, Miriel V, Bertun E (1993). Amino acid supplementation and exercise performance: proposed ergogenic value. Sports Medicine 16, 190-209
22. Lemon PWR (1991). Protein and amino acid needs of the strength athlete. International Journal of Sport Nutrition. 1, 127-145
23. Merimee TJ, Rabinowitz D, Fineberg SE (1969). Arginine-initiated release of human growth hormone. New England Journal of Medicine 280, 1434-8
24. Blomstrand E, Celsing F, Newshome EA (1988). Changes in plasma concentrations of aromatic and branch-chain amino acids during sustained exercise in man and their possible role in fatigue. Acta Physiologica Scandinavica 133, 115-21
25. Bloomstrand E, Hassmen P, Ekblom B et al (1991). Administration of branch-chain amino acids during sustained exercise – effects on performance and on plasma concentration of some amino acids. European Journal of Applied Physiology 63, 83-8
26. Bloomstrand E, Hassmen P, Newsholme E (1991). Effect of branch-chain amino acid supplementation on mental performance. Acta Physiologica Scandinavica 143, 225-6
27. Carli G, Bonifazi M, Lodi L et al (1992). Changes in exercise-induced hormone response to branched chain amino acid administration. European Journal of Applied Physiology 64, 272-7
28. Coombes J, McNaughton L (1940). The effects of branched chain amino acid supplementation on indicators of muscle damage after prolonged strenuous exercise. Medicine and Science in Sports and Exercise 27, S149 (abstract)
29. Davis JM (1995). Carbohydrates, branched-chain amino acids, and endurance, The central fatigue hypothesis. International Journal of Sport Nutrition 5, S29-38
30. Davis JM, Baily SP, Woods JA et al (1992). Effects of carbohydrate feedings on plasma free tryptophan and branched-chain amino acids during prolonged cycling. European Journal of Applied Physiology 65, 513-19
31. Gastmann UA, Lehmann MJ (1998). Overtraining and the BCAA hypothesis. Medicine and Science in Sports and Exercise 30, 1173-8
32. Hefler SK, Wildman L, Gaesser GA et al (1993). Branched-chain amino acid (BCAA) supplementation improves endurance performance in competitive cyclists. Medicine and Science in Sports and Exercise 25, S24 (abstract)
33. Kreider RB (1998). Central fatigue hypothesis and overtraining. In Kreider RB, Fry AC, OToole M (editors), Overtraining in Sport (pages 309-31). Champaign, Illinois: Human Kinetics
34. Kreider RB, Jackson CW (1994). Effects of amino acid supplementation on psychological status during and intercollegiate swim season. Medicine and Science in Sports and Exercise 26, S115 (abstract)
35. Kreider RB, Miller GW, Mitchell M et al (1992). Effects of amino acid supplementation on ultraendurance triathlon performance. In Proceedings of the I World Congress on Sport Nutrition (pages 488-536). Barcelona, Spain: Enero
36. Newsholme EA, Parry-Billings M, McAndrew M et al (1991). Biochemical mechanism to explain some characteristics of overtraining. In Brouns F (editor): Medical Sports Science, Vol. 32, Advances in Nutrition and Top Sport (pages 79-93). Basel, Germany: Karger
37. Wagenmakers AJ (1998). Muscle amino acid metabolism at rest and during exercise: role in human physiology and metabolism. In Holloszy JO (editor): Exercise and Sport Sciences Reviews (pages 287-314). Baltimore, Maryland: Williams & Wilkins
38. Kargotich S, Rowbottom DG, Keast D et al (1996). Plasma glutamine changes after high intensity exercise in elite male swimmers. Medicine and Science in Sport and Exercise 28, S133 (abstract)
39. Low SY, Taylor PM, Rennie MJ (1996). Responses of glutamine transport in cultured rat skeletal muscle to osmotically induced changes in cell volume. Journal of Physiology 492, 877-85
40. Newsholme EA, Calder PC (1997). The proposed role of glutamine in some cells of the immune system and speculative consequences for the whole animal. Nutrition 13, 728-30
41. Nieman DC, Pedersen BK (1999). Exercise and immune function. Recent developments. Sports Medicine 27, 72-80
42. Parry-Billings M, Blomstrand E, Leighton B et al (1990). Does endurance exercise impair glutamine metabolism?. Canadian Journal of Sport Science 13, 13P (abstract)
43. Parry-Billings M, Blomstrand E, McAndrew N et al (1990). A communicational link between skeletal muscle, brain and cells of the immune system. International Journal of Sports Medicine 11, S122-8
44. Parry-Billings M, Budgett R, Koutedakis K et al (1992). Plasma amino acid concentrations in the overtraining syndrome: Possible effects on the immune system. Medicine and Science in Sports and Exercise 24, 1353-8
45. Rennie MJ (1996). Glutamine metabolism and transport in skeletal muscle and heart and their clinical relevance. Journal of Nutrition 126(4), 1142S-9S
46. Rennie MJ, Tadros L, Khogali S et al (1994). Glutamine transport and its metabolic effects. Journal of Nutrirtion 124, 1503S-8S
47. Rohde T, Asp S, MacLean DA et al (1998). Competitive sustained exercise in humans, lymphokine activated killer cell activity, and glutamine–an intervention study. European Journal of Applied Physiology 78, 448-53
48. Varnier M, Leese GP, Thompson J et al (1995). Stimulatory effect of glutamine on glycogen accumulation in human skeletal muscle. American Journal of Physiology 269, E309-15
49. Birch R, Nobel D, Greenhaff P (1994). The influence of dietary creatine supplementation on performance during repeated bouts of maximal isokinetic cycling in man. European Journal of Applied Physiology 69, 268-76
50. Burke L, Pyne LD, Telford R (1996). Effect of oral creatine supplementation on single-effort sprint performance in elite swimmers. International Journal of Sports Nutrition 6, 222-33
51. Casey A, Constantin-Teodosiu D, Howell S et al (1996). Creatine ingestion favorably affects performance and muscle metabolism during maximal exercise in humans. American Journal of Physiology 271, E31-7
52. Earnest C, Snell P, Rodriguez R et al (1995). The effect of creatine monohydrate ingestion on anaerobic power indices muscular strength and body composition. Acta Physiologica Scandinavica 153, 207-9
53. Febbraio MA, Flanagan TR, Snow R et al (1995). Effect of creatine supplementation on intramuscular TCr metabolism and performance during intermittent supramaximal exercise in humans. Acta Physiologica Scandinavica 155, 387-95
54. Green AL, Hultman E, Macdonald IA et al (1996). Carbohydrate feeding augments skeletal muscle creatine accumulation during creatine supplementation in humans. American Journal of Physiology 271, E821-6
55. Green AL, Simpson EJ, Littlewood JJ et al (1996). Carbohydrate ingestion augments creatine retention during creatine feeding in humans. Acta Physiologica Scandinavica 158, 195-202
56. Greenhaff PL, Bodin K, Harris R et al (1993). The influence of oral creatine supplementation on muscle phosphocreatine resynthesis following intense contraction in man. Journal of Physiology 467, 75P (abstract)
57. Greenhaff PL, Casey A, Short A et al (1993). Influence of oral creatine supplementation on muscle torque during repeated bouts of maximal voluntary exercise in man. Clinical Science 84, 565-71
58. Greenhaff PL, Constantin-Teodosiu D, Casey A et al (1994). The effect of oral creatine supplementation on skeletal muscle ATP degradation during repeated bouts of maximal voluntary exercise in man. Journal of Physiology 476, 84P (abstract)
59. Grindstaff PD, Kreider R, Bishop R et al (1997). Effects of creatine supplementation on repetitive sprint performance and body composition in competitive swimmers. International Journal of Sports Nutrition 7, 330-46
60. Kreider R, Ferreira M, Wilson M et al (1998). Effects of creatine supplementation on body composition, strength, and sprint performance. Medicine and Science in Sports and Exercise 30, 73-82
61. Kreider RB (1998). Creatine supplementation, analysis of ergogenic value, medical safety, and concerns. Journal of Exercise Physiology On Line 1(1), www.css.edu/users/tboone2/asep/jan3.htm
62. Peeters BM, Lantz CD, Mayhew JL (1999). Effect of oral creatine monohydrate and creatine phosphate supplementation on maximal strength indices, body composition, and blood pressure. Journal of Strength and Conditioning Research (in press)
63. Peyrebrune MC, Nevill ME, Donaldson FJ et al (1998). The effects of oral creatine supplementation on performance in single and repeated sprint swimming. Journal of Sports Sciences 16, 271-9
64. Prevost MC, Nelson AG, Morris GS (1997). Creatine supplementation enhances intermittent work performance. Research Quarterly for Exercise and Sport 68, 233-40
65. Redondo D, Dowling EA, Graham BL et al (1996). The effect of oral creatine monohydrate supplementation on running velocity. International Journal of Sports Nutrition 6, 213-21
66. Smith SA, Montain SJ, Matott RP et al (1998). Creatine supplementation and age influence muscle metabolism during exercise. Journal of Applied Physiology 85, 1349-56
67. Stone MH, Sanborn K, Smith L et al (1999). Effects of in-season (5 weeks) creatine and pyruvate supplementation on anaerobic performance and body composition in American football players. International Journal of Sports Nutrition (in press)
68. Stout JR, Echerson J, Noonan D et al (1999). The effects of a supplement designed to augment creatine uptake on exercise performance and fat free mass in football players. Nutrition Research 19, 217-25
69. Terrillion KA, Kolkhorst FW, Dolgener FA et al (1997). The effect of creatine supplementation on two 700-m maximal running bouts. International Journal of Sports Nutrition 7, 138-43
70. Vanakoski J, Kosunen V, Meririnne E et al (1998). Creatine and caffeine in anaerobic and aerobic exercise, effects on physical performance and pharmacokinetic considerations. International Journal of Clinical Pharmacology and Therapeutics 36, 258-62
71. Vandenberghe K, Gillis N, Van Leemputte M et al (1996). Caffeine counteracts the ergogenic action of muscle creatine loading. Journal of Applied Physiology 80, 452-7
72. Vandenberghe K, Goris M, Van Hecke P et al (1997). Long-term creatine intake is beneficial to muscle performance during resistance training. Journal of Applied Physiology 83, 2055-63
73. Volek JS, Kraemer WJ, Bush JA et al (1997). Creatine supplementation enhances muscular performance during high-intensity resistance exercise. Journal of the American Dietetic Association 97, 765-70
74. Williams MH, Kreider RB, Branch JD (1999). Creatine: The Power Supplement. Champaign, Illinois: Human Kinetics
75. Almada A, Kreider R, Ferreira M et al (1997). Effects of calcium β -HMB supplementation with or without creatine during training on strength and sprint capacity. FASEB Journal 11, A374 (abstract)
76. Kreider R, Ferreira M, Wilson M et al (1997). Effects of calcium β -HMB supplementation with or without creatine during training on body composition alterations. FASEB Journal 11, A374 (abstract)
77. Kreider RB (1999). Dietary supplements and the promotion of muscle growth with resistance training. Sports Medicine 27, 97-110
78. Kreider RB, Ferreira M, Wilson M et al (1999). Effects of calcium β -hydroxy β -methylbutyrate (HMB) supplementation during resistance-training on markers of catabolism, body composition and strength. International Journal of Sports Medicine (in press)
79. Miller P, Sandberg L, Fuller JC (1997). The effect of intensive training and β -hydroxy-β -methylbutyrate (HMB) on the physiological response to exercise in horses. FASEB Journal 11, A1683 (abstract)
80. Nair KS, Schwartz RG, Welle S (1992). Leucine as a regulator of whole body and skeletal muscle protein metabolism in humans. American Journal of Physiology 263, E928-34
81. Nissen S, Faidley TD, Zimmerman DR et al (1992). Colostral milk fat and pig performance are enhanced by feeding the leucine metabolite β -hydroxy-β -methylbutyrate to sows. Journal of Animal Science 72, 2331-7
82. Nissen S, Panton L, Fuller J et al (1997). Effect of feeding β -hydroxy-β -methylbutyrate (HMB) on body composition and strength of women. FASEB Journal 11, A150 (abstract)
83. Nissen S, Sharp R, Ray M et al (1996). Effect of leucine metabolite beta-hydroxy-beta-methylbutyrate on muscle metabolism during resistance-exercise training. Journal of Applied Physiology 81, 2095-104
84. Van Koevering MT, Dolezal HG, Gill DR et al (1994). Effects of β hydroxy β-methylbutyrate on performance and carcass quality of feedlot steers. Journal of Animal Science 72, 1927-35
85. Vukovich MD, Stubbs NB, Bohlken RM et al (1997). The effect of dietary β -hydroxy-β -methylbutyrate (HMB) on strength gains and body composition changes in older adults. FASEB Journal 11, A376 (abstract)
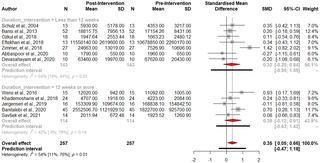
Aumento inducido por el ejercicio en el factor neurotrófico derivado del cerebro (BDNF) en personas con esclerosis múltiple: Una revisión sistemática y metaanálisis de ensayos de intervención de ejercicio
El papel del Factor Neurotrófico Derivado del Cerebro (BDNF) en la esclerosis múltiple (EM) ha...
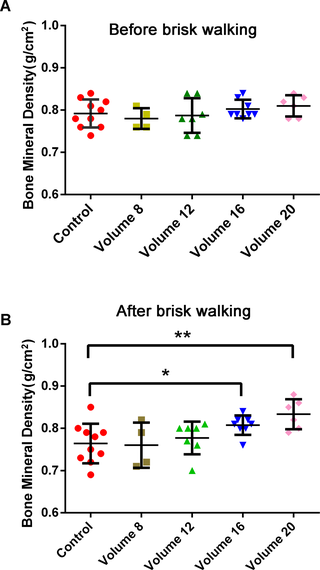
El volumen de caminata rápida es el factor clave para la mejora de la DMO en mujeres en premenopausia
Brisk Walking y Densidad Mineral Ósea: Clave para las Mujeres Premenopáusicas La osteoporosis es...
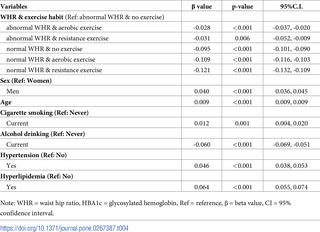
Efectos del ejercicio aeróbico y de resistencia en las concentraciones de hemoglobina glucosilada (HbA1c) en individuos taiwaneses no diabéticos según la relación cintura-cadera
El estudio analiza la relación entre los niveles de hemoglobina glucosilada (HbA1c) y la práctica...
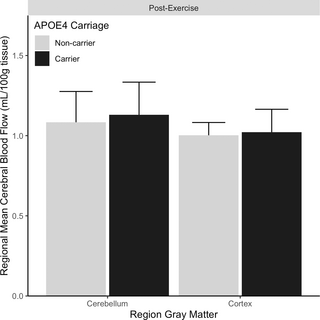
Riesgo de demencia y respuesta dinámica al ejercicio: Un ensayo clínico no randomizado
Resumen El ejercicio físico ha mostrado ser un aliado clave para la salud cerebral y la cognición...
Evaluación del proceso para el ensayo controlado aleatorizado STAMINA: Un protocolo
Evaluación del Proceso del Ensayo Controlado Aleatorizado STAMINA Introducción El ensayo STAMINA...