The Institute of Exercise and Environmental Medicine, Presbyterian Hospital of Dallas, and the University of Texas Southwestern Medical Center at Dallas, Dallas, Texas 75231.
Resumen
INTRODUCCION
Previamente, hemos demostrado que 4 semanas de aclimatación a una altura moderada (2500 m) conjuntamente con la realización de entrenamientos a baja altitud (1250 m) (HiLo) provocan resultados superiores a un entrenamiento equivalente al nivel del mar (24). Esta forma de entrenamiento en la altura produjo una mejora del 1.4% en el rendimiento de resistencia a nivel del mar en corredores universitarios y recientemente graduados (edad 20±2 años). El mecanismo que produjo la mejora del rendimiento con este enfoque parece ser doble: un incremento en la masa de glóbulos rojos resultante de la adaptación hematológica a la exposición a la altura, lo cual produjo un incremento en el máximo consumo de oxígeno (VO2 máx.), y el mantenimiento del flujo de oxígeno a nivel del mar durante el entrenamiento a baja altitud, lo cual preservó la estructura y función de los músculos esqueléticos y al a vez facilitó la mejora en el rendimiento de carrera a nivel del mar. Sin embargo, a pesar de los numerosos reportes anecdóticos respecto del éxito del entrenamiento en la altura para atletas de clase mundial, algunos reportes recientes han sugerido que el HiLo, o cualquier forma de entrenamiento en la altura, pueden no ser ventajosos para los atletas de elite, en comparación con los atletas de nivel universitario (1, 2, 14). El concepto de simorfosis, elaborado por Hoppeler y Weibel (17), establece que, para cualquier sistema, tal como la cadena respiratoria para el transporte de oxígeno, la máxima capacidad de cada parámetro es ajustada cuantitativamente para cubrir los límites estructurales y funcionales de las demandas impuestas sobre el sistema como un todo. Por lo tanto, para los “atletas de elite” del reino animal, cada paso de la vía del transporte de oxígeno desde la atmósfera a la mitocondria se ha desarrollado para realizar una función óptima y para maximizar la potencia aeróbica, permitiendo poco espacio para mejoras adaptativas adicionales. Por ello, en los atletas de elite humanos, las pequeñas mejoras a corto plazo en un paso del transporte de oxígeno pueden cubrir límites funcionales en otros pasos, minimizando los beneficios potenciales del entrenamiento en la altura. Sin embargo, los atletas de elite que viven y entrenan a nivel del mar son incapaces de desarrollar niveles similares de hemoglobina/masa de glóbulos rojos circulantes en comparación con las especies animales de “alta resistencia” que tienen la habilidad de realizar autotransfusiones por medio de la contracción esplénica (23, 25). De esta manera, la elevación de los niveles circulantes de hemoglobina tiene el mayor potencial para provocar mejoras en el rendimiento en atletas de resistencia de nivel elite. Asimismo, la interacción entre los componentes de convección y difusión del transporte de oxígeno, descritos por Wagner (35), podrían predecir el incremento en el VO2 máx. con el incremento en la hemoglobina y en la masa de glóbulos rojos circulantes. En respaldo a estos conceptos, los resultados de los estudios en donde se realizaron transfusiones (5, 10, 37) y de aquellos que han administrado eritropoyetina recombinante (3, 4) sugieren que el incremento en la masa de glóbulos rojos en si misma incrementará el VO2 máx. en todos los atletas de resistencia, sin considerar el nivel de rendimiento. Por lo tanto, el presente estudio fue diseñado para investigar el efecto del paradigma HiLo en corredores de elite quienes probablemente estén muy próximos a su máximo potencial de rendimiento respecto de los atletas previamente estudiados con esta aproximación. Este estudio fue programado para llevarse a cabo en el momento en que los atletas estuvieran en la mejor forma física de todo el año [i.e., luego de que compitieran en la temporada de verano de atletismo de pista que culminaba con el campeonato de la Asociación Nacional de Atletismo Universitario (NCAA) y el campeonato Nacional de los EE.UU. de Atletismo de Pista y Campo]. Nuestra hipótesis fue que la combinación de aclimatación a 2500 m y el entrenamiento de alta intensidad a 1250 m mejorarían el rendimiento a nivel del mar de corredores de media y larga distancia de elite.METODOS
Sujetos Veintiséis corredores de distancia (17 hombres y 9 mujeres) fueron reclutados para participar en el estudio. Se requirió que los atletas fueran de nivel nacional y que compitieran en eventos que cubrieran distancias entre los 1500 m y la maratón. Veinticuatro de los 26 atletas estaban ranqueados entre los 50 primeros de los EE.UU. para su evento en el año 1997. De estos atletas dos compitieron en los Juegos Olímpicos de 1996, y el 50% compitieron en las pruebas de clasificación para los Juegos Olímpicos de 1996. Todos los atletas, excepto cuatro, habían competido en los campeonatos de la NCAA o en el Campeonato Nacional de los EE.UU. de atletismo de pista y campo de 1997 o en ambos. Tres de estos cuatro atletas estaban intentando alcanzar la marca de calificación para el día del encuentro. Los criterios de exclusión incluyeron, la residencia en la altura (>1000m) o haber sufrido recientemente lesiones o enfermedades que evitaran el rendimiento normal durante el entrenamiento y las competencias. Los sujetos dieron su consentimiento por escrito para participar en el estudio, el cual fue aprobado por el Comité de Revisión Institucional del Centro Médico de la Universidad del Sudoeste de Texas. Protocolo El protocolo del estudio consistió en una modificación del protocolo desarrollado previamente por los autores para corredores de nivel universitario (24). En forma resumida, los atletas fueron evaluados al nivel del mar la semana antes y la semana posterior a un período de 27 días de residencia a una altura de 2500 m (ver Figura 1). El campeonato de la NCAA fue llevado a cabo a nivel del mar 3 semanas antes de la estadía en la altura, y el Campeonato Nacional de los EE.UU. de atletismo de pista y campo se realizó 1 semana antes de la estadía en la altura. Se diseñaron planes individuales de entrenamiento que fueron desarrollados por los atletas y su entrenador/a. los planes de entrenamiento fueron discutidos con los investigadores y se diseñaron conforme a un modelo de entrenamiento presentado por los investigadores (24). Se requirió que los atletas realizaran entrenamientos de alta intensidad y alta velocidad a una altitud de 1250 m. Todos los otros entrenamientos fueron llevados a cabo a altitudes de entre 1250 y 3000 m, realizándose la mayoría de los entrenamientos entre los 2000 y los 2800 metros. En un estudio piloto se ha demostrado que esta modificación del modelo HiLo, denominada “HiHiLo” (residir a moderada altitud, entrenamiento de base de baja intensidad a moderada altitud, entrenamiento fraccionado de alta intensidad a baja altitud), provoca incrementos en el VO2 máx. y mejoras en el rendimiento durante carreras de 5000 m idénticas a las observadas con el modelo original HiLo (34). Todos los atletas recibieron suplementación con hierro que se administró en forma líquida por vía oral (Feo-Sol, 9 mg de hierro elemental/ml) con la dosis ajustada en base a la concentración plasmática de ferritina (rango: 5-45 ml/día)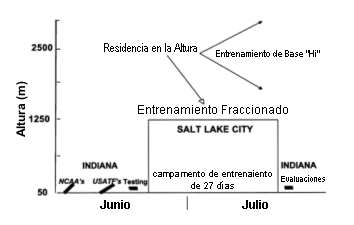 Figura 1. Calendario del experimento. NCAA, Asociación Nacional
Universitaria de Atletismo; USATF, Asociación de Atletismo de Pista y Campo de
los EE.UU.; Hi, alta altitud; Lo, baja altitud.
Figura 1. Calendario del experimento. NCAA, Asociación Nacional
Universitaria de Atletismo; USATF, Asociación de Atletismo de Pista y Campo de
los EE.UU.; Hi, alta altitud; Lo, baja altitud.
 Tabla 1. Rendimiento al nivel del mar. Los valores son presentados como
valores medios±DE. HiLo, vivir en la altura-entrenar bajo. * p≤0.05; † p<0.10 pre vs. post.
Tabla 1. Rendimiento al nivel del mar. Los valores son presentados como
valores medios±DE. HiLo, vivir en la altura-entrenar bajo. * p≤0.05; † p<0.10 pre vs. post.
RESULTADOS
Sujetos Catorce hombres (25±3 años, 179±5 cm, 63.6±5.2 kg) y ocho mujeres (24±3 años, 168±5 cm, 53.3± 4.9 kg) completaron exitosamente el protocolo para dar un total de 22 sujetos. Cuatro sujetos (tres hombres y una mujer) sufrieron lesiones (n=2) o enfermedades (n=2) durante la estadía en la altura, lo que evitó su normal desempeño durante los entrenamientos y las carreras y por lo tanto no fueron incluidos en los análisis. No se hallaron diferencias sexuales respecto de la respuesta a la estadía en la altura; por lo tanto, los datos de los hombres y de las mujeres se consideraron conjuntamente. Rendimiento El rendimiento del grupo en la prueba de 3000 m a nivel del mar mejoró significativamente luego del tratamiento HiLo (Tabla 1 y Figura 2). Los hombres y las mujeres incrementaron su rendimiento en proporciones similares, reduciendo el tiempo de la prueba en 5.8 s (95% de los límites de confianza, 1.8-1.9 s) o 1.1% (95% de los límites de confianza 0.3-0.9%). Tres atletas mejoraron su rendimiento a nivel del mar en la prueba de 3000 m en 23 s, mientras que un atleta corrió 18 s más lento.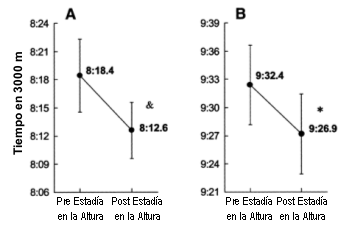 Figura 2. Cambio en el rendimiento en los corredores de elite hombres (A)
y mujeres (B) en respuesta a 4 semanas de residencia en la altura en combinación
con entrenamientos a baja altitud. *p<0.05, & p<0.10 en comparación con los valores previos a la residencia en la altura.
Figura 2. Cambio en el rendimiento en los corredores de elite hombres (A)
y mujeres (B) en respuesta a 4 semanas de residencia en la altura en combinación
con entrenamientos a baja altitud. *p<0.05, & p<0.10 en comparación con los valores previos a la residencia en la altura.
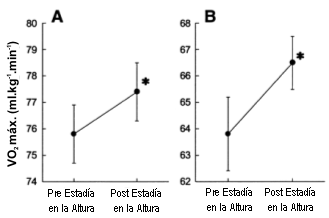 Figura 3. Cambios en el consumo máximo de oxígeno (VO2 máx.)
en los corredores de elite se sexo masculino (A) y femenino (B) en respuesta a 4
semanas de residencia en la altura en combinación con entrenamientos a baja
altitud. *p<0.05 en comparación con los valores previos a la residencia en la altura.
Figura 3. Cambios en el consumo máximo de oxígeno (VO2 máx.)
en los corredores de elite se sexo masculino (A) y femenino (B) en respuesta a 4
semanas de residencia en la altura en combinación con entrenamientos a baja
altitud. *p<0.05 en comparación con los valores previos a la residencia en la altura.
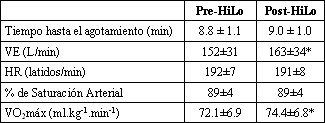 Tabla 2. VO2 máx. Los datos son presentados como valores
medios±DE; n=22 (18 hombres, 8 mujeres). VO2 máx., consumo máximo de
oxígeno; VE, ventilación minuto, HR, frecuencia cardíaca. *p≤0.05 pre vs post
HiLo.
Tabla 2. VO2 máx. Los datos son presentados como valores
medios±DE; n=22 (18 hombres, 8 mujeres). VO2 máx., consumo máximo de
oxígeno; VE, ventilación minuto, HR, frecuencia cardíaca. *p≤0.05 pre vs post
HiLo.
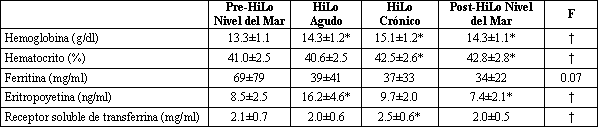 Tabla 3. Valoraciones hematológicas. Los valores son medias ± DE.
*p<0.05, diferente del valor inicial registrado a nivel del mar (test post hoc de Student-Newman-Keuls); † índice F significativo (p≤0.05) (ANOVA de una vía).
Tabla 3. Valoraciones hematológicas. Los valores son medias ± DE.
*p<0.05, diferente del valor inicial registrado a nivel del mar (test post hoc de Student-Newman-Keuls); † índice F significativo (p≤0.05) (ANOVA de una vía).
DISCUSION
El principal hallazgo de este estudio fue que, en este grupo de corredores de elite, el rendimiento en la prueba de 3000 m a nivel del mar mejoró significativamente en respuesta a 27 días de campamento en la altura utilizando el paradigma HiLo. De hecho, nueve atletas registraron su récord personal para la distancia luego del campamento HiLo, a pesar de haberse preparado para y haber competido en los eventos del campeonato nacional, antes de la estadía en la altura. El mecanismo que produjo la mejora parece ser similar al previamente descrito en estudios cuidadosamente controlados con atletas de nivel universitario (24), en donde la estimulación de la eritropoyesis derivó en un aparente incremento en el transporte de oxígeno hacia los tejidos periféricos, lo cual se evidenció por (1) un aumento de casi el doble en la concentración plasmática de eritropoyetina y una reducción del 43% en la concentración sérica de ferritina a pesar de la suplementación con hierro por vía oral durante la exposición aguda a la altura; (2) una elevación en la concentración de receptor soluble de transferrina; y (3) un incremento en la concentración de hemoglobina y en el hematocrito luego de retornar al nivel del mar, conjuntamente con la reducción de los niveles de eritropoyetina por debajo de los niveles basales originales. Limitaciones Debemos reconocer que nuestro estudio experimentó la misma limitación principal que comparten la mayoría de las investigaciones llevadas a cabo con atletas de elite: la ausencia de un grupo control que realizara un campamento de entrenamiento similar al nivel del mar. La introducción de dicho grupo control sería óptima para asegurar que los atletas no mejoraron su rendimiento meramente como resultado del entrenamiento en si mismo, sino más bien como resultado de vivir en la altura y entrenar a baja altura. Sin embargo, un gran número de evidencias sugieren que el diseño experimental empleado en este estudio fue suficiente como para explicar la mayor parte de los efectos observados. Primero, en nuestro estudio previo (24), el cual incluyó muchos años de estudios piloto, hemos determinado que para atletas de nivel universitario, eran necesarias al menos 2 semanas de entrenamiento controlado para superar el “efecto del campamento de entrenamiento”. Por ejemplo, en un estudio preliminar en donde se llevó a cabo un campamento de entrenamiento a nivel del mar, seis corredores de sexo masculino incrementaron su VO2 máx. desde 68±1.5 a 70±1.4 ml.kg-1.min-1 luego de 2 semanas de entrenamiento supervisado, pero no se observó un incremento adicional luego de otras dos semanas de entrenamiento (70±1.8 ml.kg-1.min-1) (Levine y Spray-Gundersen, observaciones no publicadas). De esta manera, para los 52 atletas hombres y mujeres estudiados en nuestros reportes previamente publicados (24, 34), luego de la fase del entrenamiento supervisado, no se produjeron incrementos significativos en el VO2 máx. con 4 semanas adicionales de entrenamiento estructurado a nivel del mar (64±0.8 a 64±0.8 ml.kg-1.min-1). Asimismo, luego de estas 6 semanas de entrenamiento a nivel del mar llevadas a cabo por atletas de nivel universitario, no se obtuvieron mejoras adicionales cuando estos participaron en un campamento de entrenamiento a nivel del mar durante 4 semanas adicionales (24). En el presente estudio, consideramos que los meses de preparación de los atletas para su participación en los campeonatos nacionales fueron al menos equivalentes a las 2 semanas de entrenamiento aplicadas a los atletas de nivel universitario varias semanas después de su temporada competitiva con el propósito de minimizar el efecto del campamento de entrenamiento. Además, la revisión del programa de entrenamiento de cada atleta para los campeonatos nacionales, sugiere que todos alcanzaron el pico de rendimiento de manera apropiada para este evento. Hasta donde pudo determinarse a partir de la inspección de las planillas de entrenamiento y de las entrevistas individuales con cada atleta durante el curso del estudio, ninguno tuvo cambios en el entrenamiento que pudieran explicar la mejora en el rendimiento. Nosotros sospechamos, pero no podemos probar, que los atletas estaban igualmente motivados para dar su mejor rendimiento en los tests realizados tanto previamente como posteriormente al campamento. Sin embargo, podemos decir que estos atletas eran extremadamente competitivos, y que nuestra impresión clínica fue que la motivación fue alta durante todas las carreras realizadas en el transcurso del estudio. Asimismo, todas las pruebas fueron corridas a un ritmo establecido por un sujeto experimentado en fijar el paso de carrera quien mantuvo la misma velocidad de carrera durante 4 de las 7.5 vueltas tanto en las pruebas pre campamento como en las pruebas post campamento. Por último, los atletas fueron estimulados a correr hasta el agotamiento durante todos los tests en cinta ergométrica, y no hubo evidencia en los datos del índice de intercambio respiratorio o de la frecuencia cardíaca que sugiriera que los atletas dieran su mayor esfuerzo durante el segundo test respecto del primero (por ejemplo, la frecuencia cardíaca máxima fue de 192 latidos/min en el primer test y de 191 latidos/min en el segundo). Tampoco podemos excluir la posibilidad de que la suplementación con hierro, llevada a cabo para asegurar la adecuada disponibilidad del mismo para la eritropoyesis durante la exposición a la altura (24) pudiera haber resultado en un incremento en la concentración de hemoglobina, independientemente del efecto provocado por la altura. Sin embargo, creemos que esta posibilidad es improbable por las siguientes razones: (1) ninguno de los atletas que participó en el presente estudio era anémico o tenía glóbulos rojos pequeños, algo característico de la anemia por deficiencia de hierro. Todos los sujetos tuvieron hematócritos y concentraciones de hemoglobina normales, y todos tenían tamaños y distribución de los glóbulos rojos normales. (2) A pesar de la suplementación con hierro por vía oral, los requerimientos de hierro durante la exposición a la altura fueron tales que las reservas de hierro de la médula ósea, medida mediante la ferritina sérica, no se incrementaron en el curso del campamento de entrenamiento. De hecho, los niveles de ferritina se redujeron en cada medición longitudinal (Tabla 3), lo que sugiere que las reservas de hierro de la médula ósea estuvieron más, en vez de menos repletadas, luego del campamento en la altura. (3) Las concentraciones basales de eritropoyetina fueron normales y bajas, lo que sugiere que no se produjo una anemia fisiológicamente significativa; la anemia es el estímulo más potente para la síntesis de eritropoyetina (20). De esta manera, la evidencia disponible, va en contra de la suposición acerca de que el simple tratamiento de la deficiencia de hierro fue el mecanismo responsable del incremento de la hemoglobina y del hematocrito en el presente estudio. Por último, debido a las limitaciones de tiempo asociadas con la utilización de atletas de elite para este estudio durante los períodos de competiciones importantes, los sujetos no fueron completamente caracterizados, ni se controlaron rigurosamente los detalles del programa de entrenamiento en la misma extensión que en nuestro trabajo previo (24). Sin embargo, para el protocolo se utilizaron los mismos modelos de entrenamiento y los parámetros clave fueron medidos en ambas poblaciones, incluyendo las concentraciones de eritropoyetina y hemoglobina, el VO2 máx., y el tiempo durante una prueba de rendimiento, lo que permitió la comparación de estos datos con los obtenidos en nuestros estudios previos. Cuando se comparan los resultados de estudios cuidadosamente controlados en donde se llevaron a cabo evaluaciones generales con corredores universitarios, con los resultados del presente estudio (Tabla 4), se observa que los resultados son notablemente similares tanto en la magnitud como en la dirección del efecto. Además, cuando se midieron parámetros similares, los resultados sugieren que el efecto fue provocado por el mismo mecanismo, i.e., un incremento en la concentración de eritropoyetina que deriva en un incremento en la concentración de hemoglobina, en un incremento en el VO2 máx. y en una mejora del rendimiento. Por lo tanto, creemos que los compromisos realizados en el presente estudio para evaluar a atletas de elite durante el período de mejor forma física no comprometió la validez de los resultados. Un Modelo Particular para Atletas de Elite Los atletas de elite del reino animal proveen un modelo particular respecto del concepto de simorfósis, por medio del cual el diseño estructural de todos los componentes que comprenden un sistema cubre cuantitativamente las demandas funcionales del sistema (17). Por ejemplo, los zorros, perros y caballos tienen una tasa de consumo de oxígeno específica para la masa ~2.5 veces mayor que las especies sedentarias del mismo tamaño corporal, tales como el agutí, la cabra y el buey. Para dichos animales, el mecanismo de este gran rango adaptativo para el consumo de oxígeno parece deberse al gran volumen mitocondrial, equiparado por un gran volumen capilar muscular y por una gran conductancia vascular en los músculos esqueléticos; una mayor concentración de hemoglobina; y un mayor volumen latido máximo. La redundancia en el sistema pulmonar de las especies no atléticas, manifestada por un exceso en la capacidad ventilatoria y de difusión, está casi eliminada en las especies atléticas, confirmando el principio de simorfósis. En otras palabras, los estudios comparativos sugieren que dichos “atletas” operan al máximo o casi al máximo del limite superior de su capacidad estructural para el transporte convectivo de oxígeno en el VO2 máx. Si este análisis es también relevante para los humanos, se podría argumentar que los atletas humanos de elite podrían tener una pequeña capacidad de adaptación para incrementar el transporte de oxígeno respecto de los atletas de menor nivel, tales como aquellos con los cuales se reportó originalmente la utilización de la aproximación HiLo. Aunque la cantidad de datos publicados en estudios que examinaron a verdaderos atletas de elite durante entrenamientos en la altura, es limitada, al menos un pequeño estudio, llevado a cabo con ciclistas de clase mundial, donde se realizaron mediciones antes y después de 31 días de estadía en la altura (2690 m) ha reportado que no se produjeron cambios en la masa de hemoglobina ni en el VO2 máx. medido a nivel del mar a pesar de la mejora en el rendimiento (14). Los autores sugirieron que el resultado se debió, al menos en parte, a la limitada reserva de adaptación en dichos atletas (14), particularmente en los pulmones, los cuales son esencialmente estáticos frente al entrenamiento. Por ejemplo, los atletas de resistencia de elite muestran limitaciones pulmonares durante el intercambio de gases a nivel del mar, mayores en magnitud y ocurrencia respecto de los individuos menos entrenados, parte de lo cual puede explicarse por las limitaciones en la ventilación (8). Sin embargo, en el presente estudio, observamos un incremento en la ventilación máxima que fue conmensurable con el incremento en el VO2 máx. No podemos determinar a partir de los datos de este experimento, si el incremento en la ventilación máxima fue una simple consecuencia del incremento en el VO2 máx. o si la causa del incremento en el VO2 máx. fue el incremento en el trabajo ventilatorio. El incremento en la ventilación durante el ejercicio y su persistencia durante un período de tiempo luego del retorno al nivel del mar, hubiera sido un resultado esperado de la aclimatación ventilatoria a las grandes altitudes y hubiera sugerido que, al menos en estos atletas, la limitación del flujo no restringió la ventilación máxima en un grado importante. Nosotros especulamos que si el incremento inicial en el VO2 máx. se requirió, al menos en parte, para respaldar el incremento en el trabajo ventilatorio provocado por la aclimatación a la altura, entonces la restauración del control respiratorio normal, a través del tiempo luego de retornar al nivel del mar hubiera permitido que el incremento en la capacidad de transporte de oxígeno sea dirigido hacia los músculos activos, proveyendo de esta manera un explicación a las observaciones citadas frecuentemente (9) acerca de que muchos atletas alcanzan sus mejores rendimientos luego de un período de reaclimatación al nivel del mar. Serán necesarios estudios adicionales para confirmar o excluir estas hipótesis.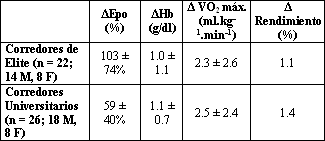 Tabla 4. Comparación de los cambios en distintas variables hematológicas
y de rendimiento observadas en atletas de elite y en atletas universitarios. Los
datos son presentados como valores medios±DE. Δ, cambio en; Epo, eritropoyetina;
M, hombres; F, mujeres. Rendimiento hace referencia al tiempo en una prueba de
3000 m (corredores de elite) o de 5000 m (corredores universitarios).
Tabla 4. Comparación de los cambios en distintas variables hematológicas
y de rendimiento observadas en atletas de elite y en atletas universitarios. Los
datos son presentados como valores medios±DE. Δ, cambio en; Epo, eritropoyetina;
M, hombres; F, mujeres. Rendimiento hace referencia al tiempo en una prueba de
3000 m (corredores de elite) o de 5000 m (corredores universitarios).
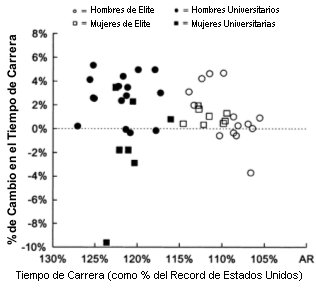 Figura 4. Comparación de atletas de diversos niveles de rendimiento y del
cambio en el rendimiento luego de 4 semanas de residencia en la altura y
entrenamientos a bajas altitudes. El tiempo de carrera representa el tiempo
registrado en la prueba inicial representado como un porcentaje del record de
Estados Unidos (AR) en el evento. El cambio en el rendimiento es el porcentaje
de cambio entre el tiempo registrado en la prueba pre campamento y el tiempo
registrado en la prueba post campamento.
Figura 4. Comparación de atletas de diversos niveles de rendimiento y del
cambio en el rendimiento luego de 4 semanas de residencia en la altura y
entrenamientos a bajas altitudes. El tiempo de carrera representa el tiempo
registrado en la prueba inicial representado como un porcentaje del record de
Estados Unidos (AR) en el evento. El cambio en el rendimiento es el porcentaje
de cambio entre el tiempo registrado en la prueba pre campamento y el tiempo
registrado en la prueba post campamento.
REFERENCIAS
1. Ashenden MJ, Gore CJ, Dobson GP, and Hahn AG (1999). Live high, train low does not change the total haemoglobin mass of male endurance athletes sleeping at a simulated altitude of 3000 m for 23 nights. Eur J Appl Physiol 80: 479484
2. Ashenden MJ, Gore CJ, Martin DT, Dobson GP, and Hahn MAG (1999). Effects of a 12-day live high, train low camp on reticulocyte production and haemoglobin mass in elite female road cyclists. Eur J Appl Physiol 80: 472478
3. Berglund B and Ekblom B (1991). Effect of recombinant human erythropoietin treatment on blood pressure and some haematological parameters in healthy men. J Intern Med 229: 125130
4. Birkeland KI, Stray-Gundersen J, Hemmersbach P, Hallen J, Haug E, and Bahr R (2000). Effect of rhEPO administration on serum levels of sTfR and cycling performance. Med Sci Sports Exerc 32: 12381243
5. Buick FJ, Gledhill N, Froese AB, Spriet L, and Meyers EC (1980). Effect of induced erythrocythemia on aerobic work capacity. J Appl Physiol 48: 636642
6. Chapman RF, Stray-Gundersen J, and Levine BD (1998). Individual variation in response to altitude training. J Appl Physiol 85: 14481456
7. Daniels J and Oldridge N (1970). The effects of alternate exposure to altitude and sea level on World class middle-distance runners. Med Sci Sports Exerc 2: 107112
8. Dempsey JA and Wagner PD (1999). Exercise-induced arterial hypoxemia. J Appl Physiol 87: 1997 2006
9. Dick FW (1992). Training at altitude in practice. Int J Sports Med 13: S203S206
10. Ekblom B, Goldbarg AN, and Gullbring B (1972). Response to exercise after blood loss and reinfusion. J Appl Physiol 33: 175180
11. Fandrey J and Jelkmann WE (1991). Interleukin-1 and tumor necrosis factor-a inhibit erythropoietin production in vitro. Ann NY Acad Sci 628: 250255
12. Faura J, Ramos J, Reynafarje C, English E, Finne P, and Finch CA (1969). Effect of altitude on erythropoiesis. Blood 33: 668 676
13. Frede S, Fandrey J, Pagel H, Hellwig T, and Jelkmann W (1997). Erythropoietin gene expression is suppressed after lipopolysaccharide or interleukin-1b injections in rats. Am J Physiol Regulatory Integrative Comp Physiol 273: R1067R1071
14. Gore CJ, Hahn A, Rice A, Bourdon P, Lawrence S, Walsh C, Stanef T, Barnes P, Parisotto R, Martin D, Pyne D, and Gore C. (1998). Altitude training at 2690 m does not increase total haemoglobin mass or sea level V˙ O2 max in world champion track cyclists. J Sci Med Sport 1: 156170
15. Gunga HC, Rocker L, Behn C, Hildebrandt W, Koralewski E, Rich I, Schobersberger W, and Kirsch K (1996). Shift working in the Chilean Andes (.3,600 m) and its influence on erythropoietin and the low-pressure system. J Appl Physiol 81: 846852
16. Hopkins WG, Hawley JA, and Burke LM (1999). Design and análisis of research on sport performance enhancement. Med Sci Sports Exerc 31: 472485
17. Hoppeler H and Weibel ER (1998). Limits for oxygen and substrate transport in mammals. J Exp Biol 201: 10511064
18. Huff RL, Lawrence JH, Siri WE, Wasserman LR, and Hennessy TG (1951). Effects of changes in altitude on hematopoietic activity. Medicine (Baltimore) 30: 197217
19. Hurtado A, Merino C, and Delgado E (1945). Influence of anoxemia on the hemopoietic activity. Arch Intern Med 75: 284323
20. Jelkmann W (1992). Erythropoietin: structure, control of production, and function. Physiol Rev 72: 449489
21. Jelkmann W, Wolff M, and Fandrey J (1990). Modulation of the production of erythropoietin by cytokines: in vitro studies and their clinical implications. Contrib Nephrol 87: 6877
22. Juvonen E, Ikkala E, Fyhrquist F, and Ruutu T (1991). Autosomal dominant erythrocytosis caused by increased sensitivity to erythropoietin. Blood 78: 30663069
23. Kraan WJ, Huisman GH, and Velthuizen J. (1978). Splenic storage volume in the unanesthetized resting beagle. Eur J Appl Physiol 38: 197206
24. Levine BD and Stray-Gundersen J (1997). Living high-training low: effect of moderate-altitude acclimatization with low-altitude training on performance. J Appl Physiol 83: 102112
25. Manohar M (1986). Right heart pressures and blood-gas tensions in ponies during exercise and laryngeal hemiplegia. Am J Physiol Heart Circ Physiol 251: H121H126
26. Ou LC, Salceda S, Schuster SJ, Dunnack LM, Brink- Johnsen T, Chen J, and Leiter JC (1998). Polycythemic responses to hypoxia: molecular and genetic mechanisms of chronic mountain sickness. J Appl Physiol 84: 12421251
27. Prchal JF and Prchal JT (1999). Molecular basis for polycythemia. Curr Opin Hematol 6: 100109
28. Reynafarje C, Lozano R, and Valdivieso J (1959). The polycythemia of high altitudes: iron metabolism and related aspects. Blood 14: 433455
29. Rice L, Ruiz W, Driscoll T, Whitley CE, Tapia R, Hachey DL, Gonzales GF, and Alfrey CP (2001). Neocytolysis on descent from altitude: a newly recognized mechanism for the control of red cell mass. Ann Intern Med 134: 652656
30. Richalet JP, Souberbielle JC, Antezana AM,Dechaux M, Le Trong JL, Bienvenu A, Daniel F, Blanchot C, and Zittoun J (1994). Control of erythropoiesis in humans during prolongad exposure to the altitude of 6,542 m. Am J Physiol Regulatory Integrative Comp Physiol 266: R756R764
31. Rusko HK, Tikkanen H, Paavolainen L, Hamalainen I, Kalliokoski K, and Puranen A (1999). Effect of living in hypoxia and training in normoxia on sea level V˙ O2 max and red cell mass (Abstract). Med Sci Sports Exerc 31: S86
32. Sanchez C, Merino C, and Figallo M (1970). Simultaneous measurement of plasma volume and cell mass in polycythemia of high altitude. J Appl Physiol 28: 775778
33. Semenza GL, Ladias JA, and Antonarakis SE (1987). An Xba I polymorphism 39 to the human erythropoietin (EPO) gene (Abstract). Nucleic Acids Res 15: 6768
34. Stray-Gundersen J and Levine BD (1997). Living high-training high and low is equivalent to living high-training low for sea level performance (Abstract). Med Sci Sports Exerc 29: S136
35. Wagner PD (1996). Determinants of maximal oxygen transport and utilization. Annu Rev Physiol 58: 215
36. Weil JV, Jamieson G, Brown DW, and Grover RF (1968). The red cell mass-arterial oxygen relationship in normal man. J Clin Invest 47: 16271639
37. Williams MH, Wesseldine S, Somma T, and Schuster R (1981). The effect of induced erythrocythemia on 5-mile treadmill run time. Med Sci Sports Exerc 13: 169175