
1Studies, Research and Sport Medicine Center, Government of Navarre, Pamplona, Spain
2Department of Physical Education, University of Las Palmas de Gran Canaria, Las Palmas de Gran Canaria, Spain
3Molecular Physiology Group, Section of Human Physiology, Department of Exercise and Sport Sciences, Copenhagen Muscle Research Center, University of Copenhagen, Copenhagen, Denmark
4Department of Physiological Sciences I, Institut d’Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS), University of Barcelona, Barcelona, Spain
Resumen
INTRODUCCIÓN
El ejercicio de fuerza regular es un componente esencial de los programas de intervención efectiva diseñados para mejorar la fuerza en atletas y adultos con enfermedades crónicas e invalideces [1], [2]. La respuesta al programa del entrenamiento de la fuerza depende finalmente de las adaptaciones metabólicas y morfológicas de las múltiples funciones celulares que dependen de una gran magnitud de los cambios de una red compleja de señalización que está implicada durante cada sesión de entrenamiento en respuesta a la actividad contráctil [3], [4].
Aunque los cambios en los metabolitos musculares y la potencia durante ejercicios como el pedaleo [5], [6], la carrera [7] o la extensión de rodillas isométrica [8], en forma exhaustiva y no exhaustiva fuertemente intermitente, están bien caracterizados, poco se sabe de la utilización del sustrato y la demanda metabólica durante series consecutivas hasta al agotamiento comparadas con no agotamiento en un ejercicio de fuerza dinámico de alta intensidad. La respuesta adaptativa al entrenamiento de la fuerza puede ser diferente cuando el entrenamiento que se lleva a cabo hasta al fallo, se lo compara a cuando no se lo lleva hasta al fallo, ya que diferentes grados de fatiga y acumulación del metabolito muscular se producen por el entrenamiento [9]. El propósito de este estudio fue, por lo tanto, investigar la influencia del número de repeticiones por serie (llevando hasta al fallo o no) sobre los cambios en el músculo y metabolitos en sangre y la potencia durante el ejercicio del press de piernas bilateral de alta intensidad realizado con la misma carga inicial (~83% 1MR) en hombres entrenados jóvenes, examinando la potencia simultáneamente y la fatiga desarrollada a lo largo del ejercicio. Estas dos sesiones de ejercicio se usan tradicionalmente por alcanzar metas específicas de entrenamiento. Así, el ejercicio "que lleva al fallo" se caracteriza por una progresiva disminución en la carga y potencia a lo largo de las series repetidas y se usa principalmente para aumentar la fuerza muscular y la hipertrofia [10], [11]. El ejercicio ‘que no lleva al fallo" se caracteriza por el mantenimiento de la carga y la potencia promedio a lo largo de las series y se usa principalmente para optimizar el desarrollo de la potencia muscular [12]. Para el conocimiento del autor, ningún estudio ha analizado los cambios en el músculo y metabolitos en sangre durante el ejercicio dinámico de alta intensidad caracterizado por el mantenimiento de la potencia a lo largo de las series. Analizando diferentes condiciones relacionadas a los cambios en la potencia, pueden dar algunas pistas a la comprensión de los mecanismos por los cuales el proceso de contracción muscular intenta mantener una función adecuada durante el ejercicio de fuerza dinámico [13]. Un segundo propósito del estudio fue examinar la relación entre el estado metabólico del músculo y los cambios en la potencia. Este tipo de examinación puede mejorar el conocimiento de los factores que limitan la fatiga durante el ejercicio del press de piernas, y así dar una indicación de la regulación de las vías metabólicas y de cómo los mecanismos anaeróbicos están relacionados durante el ejercicio de fuerza dinámico en el hombre.
MATERIALES Y MÉTODOS
Sujetos
Seis voluntarios varones sanos participaron en el estudio. Su promedio (±SD) de edad, altura, masa corporal, índice de masa corporal, consumo máximo de oxígeno estimado (VO2máx) en cicloergómetro y fuerza máxima (1MR) durante el ejercicio del press de piernas bilateral, era 34±6 años, 179±5 cm, 74.5±7.2 kg, 23.3±1.7 Kg·m-2, 57.1±4.9 ml·kg-1·min-1 y 199±43 kg, respectivamente. Todos eran atletas entrenados, principalmente en eventos de resistencia, pero ninguno se entrenaba para la competición. El porcentaje promedio de las fibras de contracción lenta (ST) para estos sujetos era del 65±12%. Los sujetos estaban completamente informados del propósito, naturaleza, detalles prácticos y posibles riesgos asociados con el experimento, así como el derecho para terminar la participación a voluntad, antes de que ellos dieran su consentimiento informado por escrito para participar. Una examinación médica también fue llevada a cabo por un médico. El presente estudio es parte de un proyecto que ha examinado los efectos metabólicos, neurales y del entrenamiento del ejercicio del press de piernas y ha sido aceptado por el Comité de Revisión Institucional del Instituto Navarro del Deporte, según la Declaración de Helsinki.
Protocolo experimental y diseño
Este estudio fue diseñado para examinar la influencia del número de repeticiones por serie (llevando vs no llevando al fallo) en los cambios en los metabolitos musculares y la potencia durante el ejercicio del press de piernas bilateral de alta intensidad. Para eliminar cualquier posible efecto de factores desconocidos, distintas variables como la carga inicial y el número total de repeticiones fueron controlados igualando sus valores entre las dos sesiones de ejercicio. Cada sujeto participó en días separados dos experimentos en los que realizaron 50 repeticiones con la misma carga inicial. Esta carga inicial (es decir, 154±31 Kg o el 83% 1MR, la carga más pesada que podría usarse una sólo una vez con la técnica correcta) fue el peso máximo que era posible completar 10 repeticiones hasta el fallo (10RM) en el ejercicio del press de piernas. El fallo se indicó como la incapacidad para completar la próxima repetición. En un día experimental (protocolo ‘que llevaba hasta el fallo’) los sujetos realizaron 5 series de 10 repeticiones hasta el fallo (10REP), separadas por 2 min de pausa entre cada serie. En este día experimental los sujetos pudieron terminar todas las 10 repeticiones a la carga inicial durante la primera serie. A veces, sin embargo, los sujetos no podían levantar la carga inicial durante las siguientes series debido a la fatiga. En el momento en que los sujetos no podían levantar la carga, se disminuía por 15 Kg, permitiéndoles así completar la prueba (50 repeticiones). En el otro día experimental (protocolo ‘que no llevaba hasta el fallo’), los sujetos realizaron 10 series de 5 repeticiones (5REP) sin llegar al fallo con la misma carga inicial como que la de (10REP), separadas por 2 minutos de pausa entre cada serie. Cinco repeticiones por serie fueron escogidas durante este día experimental de ‘no fallo’ porque éste es el número máximo de repeticiones a la que la potencia máxima puede mantenerse o ligeramente (10-15%) menor durante una serie de 10 repeticiones hasta el fallo en el ejercicio de extensión de piernas [14]. En 5REP todos los sujetos pudieron terminar el protocolo entero con la carga inicial y mantener la potencia promedio a lo largo de las series repetidas. En 10REP, sin embargo, todos los sujetos disminuyeron la potencia promedio a lo largo de las series.
Período de intervención
Todos los sujetos participaron en los dos experimentos en orden aleatorio. Los experimentos se llevaron a cabo al mismo momento del día, separados uno a dos meses. Ningún cambio se observó en los sujetos en la fuerza del press de piernas máxima (1MR) entre el primer día experimental (194±25 Kg) y el segundo día experimental (185±32 Kg). Para evitar alguna perturbación de los sujetos, se les dijo que registren su dieta normal por 48 horas antes del primer día experimental y repetir lo mismo de su dieta antes del segundo día experimental.
Tests preliminares
Algunas sesiones de pre-test tuvieron lugar durante las 3 semanas que precedieron a los experimentos. Primero, los sujetos fueron acostumbrados de antemano con los procedimientos del análisis experimental aproximadamente 2 semanas. Segundo, dos semanas antes del primer experimento, los sujetos participaron en un día de evaluación de control donde las verificaciones de la carga de fuerza para el valor de 1MR fueron determinadas en la máquina de ejercicio del press de piernas. Luego, después de al menos 10 minutos de descanso, los sujetos realizaron una serie repetitiva máxima hasta el fallo con la carga que teóricamente puede producir 10 repeticiones hasta la fatiga (~85% de 1MR). Si el número de repeticiones hasta el fallo era igual a 10, la carga era definida como 10RM y usada durante los tests principales experimentales. Si el número de repeticiones hasta el fallo era diferente de 10, varias pruebas de una serie repetitiva máxima hasta el fallo se realizaban en días diferentes con cargas más bajas o más altas durante las sesiones de prueba subsecuentes para determinar la carga que conducía hasta el fallo en exactamente 10 repeticiones. Tercero, el consumo máximo de oxígeno (VO2máx) de cada sujeto fue estimado [15] en un día separado usando un test incremental continuo hasta el agotamiento en un cicloergómetro con carga por fricción (Monark Ergomedic 818E, Varberg, Suecia). La primera carga de trabajo (60 W) era lo suficientemente alta para asegurar que el agotamiento ocurra dentro de los 8-14 minutos, siendo la carga incrementada por 30 W al final de cada minuto. La frecuencia cardíaca fue supervisada continuamente a lo largo del test (15 segundos) con un cardiotacómetro (Sportester Polar, Kempele, Finlandia). La potencia promedio en el agotamiento fue de 347±27 W.
Tests principales
En la mañana del experimento, los sujetos arribaron después de un desayuno ligero y un período de ayuno de 2 horas. Al llegar los sujetos al laboratorio, descansaron en una cama durante 20 minutos para que pudieran hacerse pequeñas incisiones bajo anestesia local (2 ml, 1% lidocaína) en la piel y fascia del músculo vasto externo de una pierna. Los sujetos luego completaron un período de precalentamiento que consistió en una serie de 5 repeticiones al 50%, tres a cuatro repeticiones al 75% y 1 repetición al 90% de la fuerza máxima del press de piernas bilateral (1MR). Se hicieron tres a cuatro intentos subsecuentes para determinar el valor de 1MR. El período de descanso entre los intentos máximos siempre era de 2 minutos. Después de 10 minutos de descanso, una biopsia muscular (la biopsia inicial) se tomó de la región media del músculo vasto externo (15 cm sobre la rótula y aproximadamente 3 cm por debajo de la entrada a través de la fascia) y después de una muestra de sangre arterializada fue tomada del lóbulo, previamente hiperemizado con Finalgon® (Boehringer Ingelheim, Alemania). Luego, ellos realizaron hasta el fallo 5 series de 10 repeticiones (10REP) o 10 series de 5 repeticiones sin llegar al fallo (5REP) con la carga máxima posible lograr 10 repeticiones durante la primera serie (10RM). Se les dijo a los sujetos que siempre intenten desplazar el peso tan rápido como fuera posible. La duración de cada repetición descompuesta en sus componentes concéntrico y excéntrico fue registrada. Las repeticiones eran interpuestas por pausas de ~1 segundo para impedir la mejora del rendimiento del ciclo de estiramiento-acortamiento. La potencia de cada repetición fue monitoreada continuamente y medida durante la fase de acción concéntrica del press de piernas. Inmediatamente (dentro de 5-10 segundos) después de la última repetición de la primera serie e inmediatamente después de la última repetición de la última serie, fueron tomadas las biopsias musculares en todos los sujetos. Las muestras de sangre arterializada adicionales fueron tomadas después de 16 y 45 minutos de recuperación para determinar la concentración sanguínea de ácido úrico post-ejercicio. A todos los participantes se les dio un estímulo verbal muy motivador y fuerte para que alentarlos a que realicen cada repetición al máximo y tan rápidamente como fuera posible. Los sujetos permanecieron en ayunas durante todos los ensayos.
Equipamiento
El estudio se realizó en una máquina de resistencia variable de extensión de piernas bilateral horizontal (es decir, la acción del press de piernas en una posición sentada; Technogym, Gambettola, Italia). La posición sentada fue ajustada para minimizar el desplazamiento entre la espalda inferior y el respaldo durante el esfuerzo de fuerza muscular individual y, por lo tanto, evitar cambios de la postura. Se les dijo a los sujetos que pongan sus pies en la misma posición en la plataforma de fuerza. La máquina de ejercicio incorporó cuatro transductores de fuerza en una plataforma de apoyo localizados debajo de los pies del sujeto. El extensiómetro registraba la fuerza aplicada (N) a una exactitud de 1 Newton en 1000 Hz. La plataforma de fuerza y el apoyo del press de piernas permanecían estacionarios a lo largo del levantamiento, mientras el cuerpo movía por fuera de los pies. En suma, un encoder rotatorio (Computer Optical Products Inc, California, EE.UU.) se ató a los platos de peso para registrar la posición y la dirección del desplazamiento a una exactitud de 0.2 mm en 1000 Hz. Un software personalizado fue usado para calcular la potencia (producto inmediato de velocidad del desplazamiento y la fuerza aplicada) por repetición. Después del final del ejercicio, los resultados fueron integrados en intervalos de 1-ms. La máxima integral de 10-ms de la fuerza aplicada y la velocidad del desplazamiento durante cada repetición es definida como la "potencia pico". La integral promedio de 10-ms de la fuerza aplicada y la velocidad del desplazamiento durante el tiempo de la contracción concéntrica total de cada repetición es definida como la "potencia promedia".
Muestras musculares
Las biopsias musculares se tomaron como fue descrito por Bergstrom [16] de la pierna derecha en cada ocasión. Las muestras musculares fueron inmediatamente congeladas (en 5-10 segundos) en nitrógeno líquido y guardadas a -80°C para el análisis histoquímico y pruebas de metabolitos subsecuente, después se las libró de grasa visible y de tejido conectivo. [16].
Análisis
La fosfocreatina (PCr), creatina (Cr) y lactato musculares, se analizaron por análisis fluorométrico [17]. Los nucleótidos de adenina y el monofosfato de inosina (IMP) del músculo esquelético se analizaron por cromatografía líquida de alto rendimiento (high-performance liquid chromatography, HPLC) [18]. Las concentraciones de los metabolitos musculares se expresan todas en mmol·Kg-1 de músculo húmedo. En suma, una porción de la biopsia tomada antes de la primera serie fue congelada y separada, después usada para el seccionamiento cruzado serial (10 µm) y marcada para la ATPasa miofibrilar luego de la preincubación alcalina y ácida [19] para la clasificación de las fibras en el tipo de contracción lenta (ST) y de contracción rápida (FT).
Cálculos
La carga de energía celular, una medida de la magnitud a la cual el pool de nucleótidos de adenina total de la célula (ATP, ADP y AMP) se fosforila, fue estimada usando la ecuación siguiente:
Carga energética = ([ATP]+0.5[ADP])/([ATP]+[ADP]+[AMP]
Muestras de sangre
Se obtuvieron muestras de sangre capilares para la determinación de las concentraciones del ácido úrico en reposo desde un lóbulo hiperemizado y 16 y 45 minutos después del final del protocolo del ejercicio. Según las instrucciones del fabricante, después de limpiar y punzar una única muestra capilar de 28.5-31.5-KL se tomó y se puso encima de la tira (ácido úrico Reflotron) para un análisis de fotometría reflectante automática (Reflotron; Boehringer Mannheim, Mannheim, Alemania) dentro de los primeros 2-3 minutos después de obtener la muestra. El analizador fue calibrado (Reflotron Check) antes del análisis de las muestras capilares de cada sujeto.
Estadística
Se usaron métodos estadísticos estándar para el cálculo de la media y la desviación estándar (SD). El t-test de Student apareado se usó para las comparaciones de los valores analíticos durante las dos condiciones experimentales diferentes en este estudio, mientras que el análisis de sentido único de variación para las medidas repetidas fue usado para examinar las diferencias a lo largo del tiempo en los índices del rendimiento y las concentraciones de metabolitos. Cuando un valor F significativo era alcanzado (P <0.05), las medias fueron comparadas usando un test post hoc LSD. Para propósitos de comparación, la potencia para las 5 repeticiones segundas de cada serie se comparó con la de las primeras 5 repeticiones. El coeficiente de determinación (R2) fue usado para determinar las asociaciones entre las variables tomando ambas condiciones de ejercicio en conjunto. Las regresiones lineales y no lineales fueron determinadas usando una forma polinómica lineal o de segundo grado. Una forma polinómica de segundo grado fue aceptada si producía una reducción significativa en la variación del error comparado con la solución lineal. La significancia estadística se aceptó al nivel de P <0.05.
RESULTADOS
Producción de fuerza y potencia
Durante 5REP todos los sujetos pudieron completar todas las repeticiones con la carga inicialmente asignada (154±31 Kg; 83±8% de 1MR). Durante 10REP, sin embargo, la mayoría de los participantes fue incapaz de completar todas las repeticiones con esta carga inicial, debido al fallo. La carga tuvo que ser reducida por 7.2±3.8% después de 27±16 repeticiones y fue progresivamente reducida, alcanzando el 85±12% (P <0.05) de la carga inicial en la última repetición. La carga promedio durante las 50 repeticiones de 10REP fue un 6.1±6.3% más baja (P <0.05) que durante 5REP.
La potencia pico promedio durante las primeras 5 repeticiones de la primera serie fue similar en ambas condiciones experimentales (Fig. 1). Durante cada serie de 5REP el valor más alto de potencia pico promedio se alcanzó durante la segunda o tercera repetición y después de esto, la potencia pico promedio disminuyó progresivamente (7-20%, P <0.05) entre la segunda y la quinta repetición a lo largo de cada serie. Durante cada serie de 10REP el valor más alto de potencia pico promedio también se alcanzó durante la segunda o tercera repetición y después de esto, la potencia pico promedio disminuyó agudamente (35-45%, P <0.01) entre la segunda y la décima repetición a lo largo de cada serie. Durante 10REP la magnitud de la disminución en la producción de la potencia pico promedio entre las primeras 5 repeticiones de la primera serie y las 5 repeticiones últimas de la serie última fue del 33±19% (de 821±209 a 569±159 W; P<0.05). En contraste, durante 5REP la producción de potencia pico de promedio en los grupos de 5 repeticiones se mantuvo entre las series. Cuando se compararon ambos tipos de ejercicio en grupos de 5 repeticiones, la potencia pico promedio de las repeticiones 6 a 50 era un 28±12% más baja (P <0.01) durante 10REP que durante 5REP. En ambas condiciones experimentales, los cambios de la potencia media, en promedio, se asemejaban a los de la producción de la potencia pico.
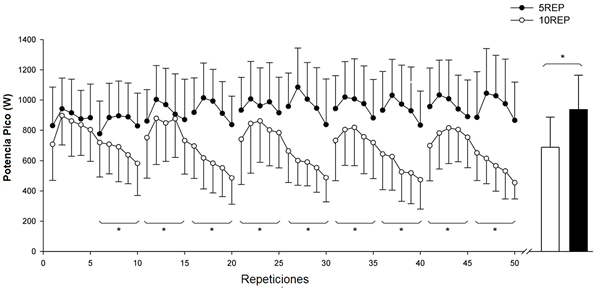
Figura 1. Perfiles de la producción de potencia pico (promedio para n=6 sujetos) para cada ejercicio durante las dos condiciones experimentales: cuando el ejercicio era 5 series de 10 repeticiones al fallo (10REP; círculos blancos), y cuando el ejercicio era 10 series de 5 repeticiones sin llegar al fallo (5REP; círculos negros). Las barras representan la media de la producción de la potencia pico a lo largo de las 50 repeticiones para 10REP y 5REP. * diferencia significativa (P <0.05) entre 10REP y 5REP (agrupados de 5 a 5 repeticiones). Los valores son promedios ±SD.
La Tabla 1 muestra las concentraciones de los metabolitos musculares antes de la primera serie (biopsia inicial) e inmediatamente después de la primera y de la última serie durante ambos días experimentales. Las concentraciones iniciales de los metabolitos estuvieron dentro del rango normal para el músculo esquelético humano. Al final de las 10REP, las reservas de PCr casi se depletaron (85% de caída, P <0.05), aunque el ATP (21%), la carga de energía (4%), y la cantidad total de pool de los nucleótidos de adenina (ATP + ADP + el AMP) (20%) estuvieron reducidos (P <0.05). Simultáneo a esto, el IMP y el lactato aumentaron. En contraste a 10REP, 5REP produjo notablemente una disminución inferior de la PCr del músculo (~15% de caída, P <0.05) y el ATP, IMP, cargo de energía y la cantidad total de concentraciones del pool de nucleótidos de adenina del músculo estuvieron inalterados, mientras que el lactato muscular sólo se elevó ligeramente sobre los niveles iniciales. Los cambios observados en los niveles de ATP del músculo, del pool de nucleótidos de adenina total, del IMP, de la PCr y del lactato al final del ejercicio, fueron significativamente superiores (P <0.01-0.05) durante 10REP comparado con 5REP.
Comparado con los valores iniciales, al final del ejercicio las proporciones de ATP/ADP calculadas disminuyeron por 9% (P <0.05) en 5REP y por 30% (P <0.01) en 10REP (Tabla 2). La disminución de la proporción de ATP/ADP en 10REPS fue más alta que la disminución correspondiente observada en 5REP. La proporción de ATP/AMP no cambió al final de 5REP pero disminuyó (P <0.05) por 34% al final de 10REP. Similarmente, la proporción de ATP/IMP no cambió después de 5REP pero mostró una disminución pronunciada de 1049 a 13.4 (P <0.05) al final de 10REP. Ningún cambio ocurrió en la proporción de ADP/AMP a lo largo de los ejercicios.
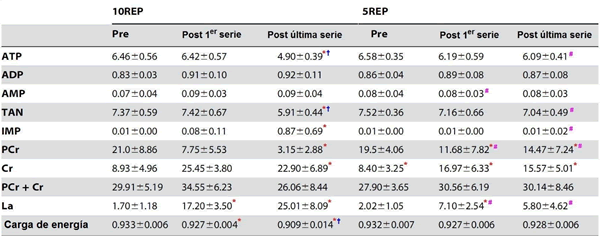
Tabla 1. Efectos del ejercicio del press de piernas sobre los nucleótidos de adenina del músculo, IMP, PCr, Cr, lactato y carga de energía antes de la primera serie, después de la última repetición de la primera serie y después de la última repetición de la última serie, durante los ejercicios de 10REP y de 5REP.
Los valores están expresados como promedios ±SD en mmol·kg-1 peso músculo húmedo, excepto cargo de energía; n = 426. TAN, total nucleótidos de adenina (ATP + ADP + AMP); IMP, Inosina 5’-monofosfato; PCr, Fosfocreatina; Cr, Creatina; La, Lactato. Para los cálculos de la carga de energía ver MÉTODOS.
* diferencia significativa (P <0.05) con el valor de pre-ejercicio.
† diferencia significativa (P <0.05) con el valor post de la primera serie.
# diferencia significativa (P <0.01-0.05) con el ejercicio 10REP.
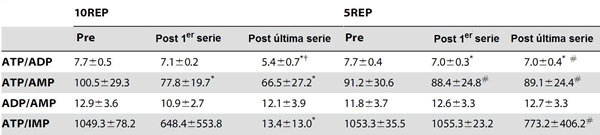
Tabla 2. Efectos del ejercicio del press de piernas sobre las proporciones de los metabolitos nucleótidos antes de la primera serie y durante los protocolos 10REP y 5REP.
Los valores están expresados en promedios ±SD.
* diferencia significativa (P <0.05) con el valor de pre-ejercicio.
† diferencia significativa (P <0.05) con el valor post de la primera serie.
# diferencia significativa (P <0.01-0.05) con el ejercicio 10REP.
Ácido úrico en sangre
La concentración sanguínea de ácido úrico máxima durante la recuperación después de la última serie de 10REP (351±96 µmol·l-1) fue un 19% más alta (P <0.05) que los valores iniciales (276±52 µmol·l-1). En 5REP, la concentración sanguínea de ácido úrico máxima después del ejercicio (292±74 µmol·l-1) permaneció inalterada de los valores de inicio (286±701 µmol·l-1). La concentración sanguínea de ácido úrico post-ejercicio durante 10REP fue más alta (P <0.05) que durante 5REP.
Relaciones entre los metabolites musculares
Para examinar las relaciones entre las variables, ambas condiciones de ejercicio fueron tomadas en conjunto. Una significativa relación curvilínea negativa fue observada entre el lactato muscular después de la primera y la última serie y los niveles correspondientes de PCr del músculo (en % de los niveles iniciales) (R2 = 0.46, P <0.05) (Figura 2A). Similarmente, una relación curvilínea significativa se observó entre la disminución relativa en los niveles de PCr del músculo (en % de los niveles iniciales) y los niveles de IMP del músculo (R2 = 0.44, P <0.01) (Figura 2B). De estas relaciones curvilíneas puede estimarse que un 60% de disminución en la PCr por debajo de los valores de reposo, es requerido para provocar una acumulación de IMP muscular.
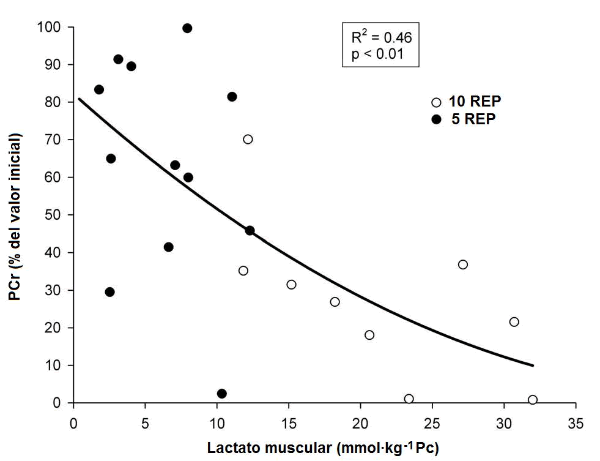
Figura 2. Relación individual entre las concentraciones del lactato muscular y las concentraciones de la PCr (expresadas en el porcentaje del valor inicial) (2A) y entre las disminuciones de la PCr (expresadas en el porcentaje de valor inicial) y las concentraciones de IMP del músculo (2B), durante 10REP (círculos blancos), y 5REP (círculos negros).
Relaciones entre los metabolitos musculares y los cambios en la potencia
Una significativa correlación negativa lineal se observó entre los cambios promedio en la potencia pico observada durante las dos últimas repeticiones (expresados en el porcentaje de los valores de las dos repeticiones iniciales) y las disminuciones en los niveles de ATP (expresadas en el porcentaje del valor inicial) (R2 = 0.59, P <0.01) (Figura 3). Es más, una significativa correlación negativa curvilínea se observó entre los cambios de la potencia pico promedio observados durante las dos últimas repeticiones de la primera y última series (expresados en el porcentaje de los valores de las dos repeticiones iniciales) y los niveles correspondientes de lactato muscular (R2 = 0.64, P <0.01) (Figura 4). La naturaleza curvilínea de la curva parece indicar que cuando los niveles del lactato muscular no exceden el límite superior de 10-15 mmol·Kg-1 de músculo húmedo, la potencia cambia poco de los valores máximos. Sin embargo, cuando los valores del lactato muscular exceden este nivel superior, la potencia disminuye en gran medida.
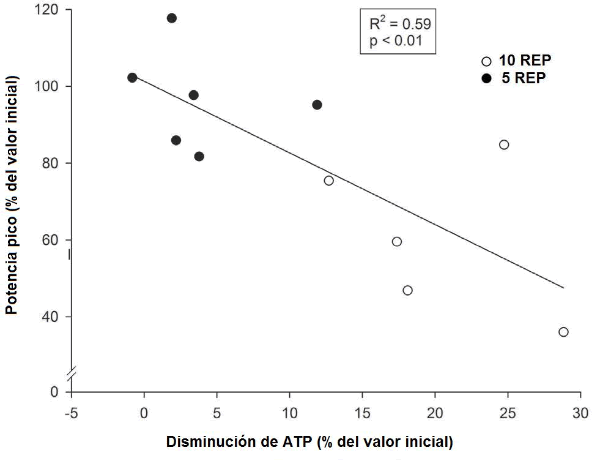
Figura 3. Relaciones individuales entre los cambios relativos de la potencia pico promedio (expresados en el porcentaje del valor inicial) entre la primera y las dos últimas repeticiones, y las disminuciones de ATP (expresadas en el porcentaje del valor inicial), durante el ejercicio 10REP (círculos blancos), y el ejercicio 5REP (círculos negros).
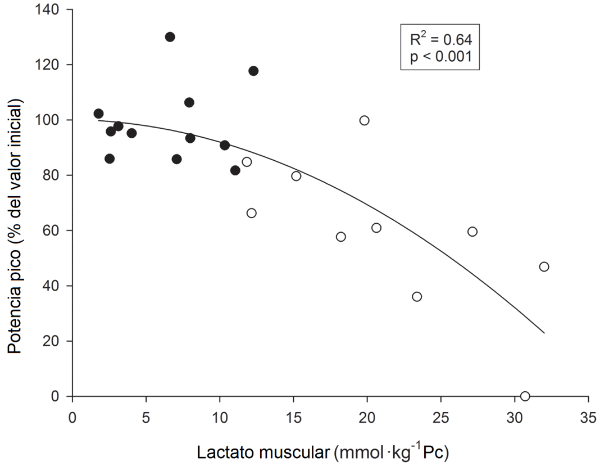
Figura 4. Relaciones individuales entre los cambios relativos de la potencia pico promedio (expresados en el porcentaje del valor inicial) entre la primera y las dos últimas repeticiones de la primera serie y entre las primeras y últimas dos repeticiones del ejercicio, y las concentraciones del lactato muscular, durante el ejercicio 10REP (círculos blancos), y el ejercicio 5REP (círculos negros).
DISCUSIÓN
Metabolismo muscular
El ejercicio 10REP de press de piernas exigió un esfuerzo máximo de los sujetos, como quedó reflejado por la marcada disminución en la potencia que tuvo lugar en cada serie durante las 5 últimas repeticiones. El uso de este modelo de 10REP causó un estado de deficiencia de energía y una disminución en el potencial de los fosfatos, resultando en un vaciamiento casi completo de las reservas de PCr, una reducción significativa en el ATP (21%) y el tamaño del pool de los nucleótidos de adenina total (TAN, total adenine nucleotide), y en marcados aumentos en el lactato y acumulación del IMP en el músculo, así como niveles altos de ácido úrico en la sangre. Los elevados niveles del lactato muscular indican que la glucólisis anaeróbica es extensivamente activada durante este tipo de ejercicio. El mayor incremento de IMP muscular se origina de la degradación del ATP y parece reflejar un fallo de la resíntesis del ATP para coincidir con la tasa de hidrólisis del ATP [13]. La elevada acumulación plasmática de ácido úrico después de las 10RP indica que el IMP no volvió a reaminarse a AMP después del ejercicio, y que, en cambio, hubo una desfosforilación del IMP a inosina, y por consiguiente, a hipoxantina y ácido úrico [20]. El resultado neto pudo haber sido una pérdida excesiva de purinas del músculo [20] que podría requerir el reemplazo por el ciclo de nucleótidos de purina o la síntesis de nuevo [4]. La magnitud de la producción de energía anaeróbica y la caída de la potencia pico con 10REP es más alto que los cambios encontrados previamente en culturistas usuarios de esteroides anabólicos luego de un régimen de ejercicio que comprendía 5 series de 10MR cada una de sentadillas frontales, sentadillas con barra atrás, press de piernas y extensiones de rodillas [21]. Sin embargo, es cualitativamente y cuantitativamente similar a lo que ha sido reportado previamente por otros durante y después de pedaleo de alta intensidad intermitente [5], [6], ejercicio de rodilla isométrico [8], o entrenamiento isocinético [22] que llevaban al agotamiento entre 6 y 60 segundos. Estos cambios también han sido asociados con grandes reducciones en el glucógeno muscular [6] y ATP [6], [22], particularmente en las fibras de tipo II [23], con cambios más pronunciados que son localizados en las fibras de tipo IIx [24], mientras que el pH en sangre y el pH muscular pueden alcanzar valores tan bajos como 7.1 [5] y 6.6 [25] respectivamente. Sin embargo, el protocolo experimental en los estudios previos no era igual que en el presente estudio, ya que se usaron acciones isoinerciales de contracciones repetitivas en el press de piernas. Así, parece ser que 10REP causa una marcada ruptura en el balance de energía en el músculo.
En contraste a las 10 repeticiones con una carga de 10MR (10REP), el balance de energía se mantuvo durante 5 repeticiones con una carga de 10MR (5REP), a pesar de la acumulación de las series. De esta manera, comparado con 10REP, 5REP resultaron en una notable disminución en el contenido de la PC en el músculo (~15% vs 80% de caída) con sólo aumentos modestos en el lactato muscular y ningún cambio mensurable en los niveles musculares de ATP e IMP y en los niveles sanguíneos de ácido úrico. Estos cambios menores de los metabolitos observados en 5REP, a pesar de una alta tasa de recambio de ATP, indican que la síntesis de ATP coincidió con la tasa de utilización de ATP y esa homeostasis celular fue mantenida, demostrando, por consiguiente, que la tasa de desaminación del AMP fue baja [13]. Por lo tanto, la producción de la potencia pico sólo se mantuvo durante el ejercicio de 5REP. La ausencia de aumento del ácido úrico sanguíneo durante 5REP está en línea con el hallazgo de niveles de ATP inalterados. Estos resultados indican que reduciendo el número de repeticiones por serie al 50%, se mantiene la potencia y el balance de energía en el músculo a lo largo de las series.
Los cambios en las proporciones de ATP/ADP y de ATP/AMP durante el proceso de la contracción han sido ampliamente estudiados porque los niveles relativos de los nucleótidos de adenina son los reguladores metabólicos más importantes para el mantenimiento de las funciones celulares adecuadas que la concentración absoluta de ATP [13]. Al final de 5REP, la proporción de ATP/ADP calculada disminuyó un 9% comparado con los valores iniciales, y fue coincidente con los pocos cambios en la carga de energía, el AMP, y las proporciones de ATP/AMP y ATP/IMP, con un 7-20% de disminución en la potencia pico promedio entre la segunda y la quinta repetición de cada serie, y con disminuciones moderadas (30%) en los niveles de la PCr. La dicotomía entre los cambios en la proporción de ATP/ADP y los cambios en la proporción de ATP/AMP durante el ejercicio no carece de precedente. De esta manera, se han observado aumentos en el contenido de ADP libre muscular estimado, sin ningún cambio en el AMP libre estimado, durante un sprint de pedaleo isocinético de 15 segundos [26] o después de 10 minutos de pedalear al 65% del consumo máximo de oxígeno [27], indicando que un aumento inicial ligero en la disponibilidad de ADP no desplaza a la adenilato kinasa favoreciendo la formación de AMP (ADP + ADP ⟷ AMP + ATP), cuando la disminución en la producción de la potencia y los cambios en la PCr del músculo o niveles del Pi son moderados. La actividad de la adenilato kinasa todavía fue aún menor luego de 5REP, probablemente porque la desfosforilación de ADP a AMP es tamponada por la PCr [23]. En esta situación, la proteína kinasa AMP-activada (la AMPK) sólo puede activarse ligeramente por la mayor disponibilidad de ADP [28]. Al final de 10REP, sin embargo, una disminución mayor (30%) en la proporción de ATP/ADP, superior a la vista después de 5 REP, fue acompañada por las disminuciones en las proporciones de ATP/AMP y de ATP/IMP, en paralelo con mayores disminuciones (33%) en la potencia y con los niveles de la PCr casi depletados. Las menores proporciones de ATP/AMP y de ATP/IMP indican que, como opuesto a 5REP, durante 10REP la adenilato kinasa y la AMP deaminasa fueron significativamente activadas. Los niveles muy bajos de PCr y la activación de la adenilato kinasa podrían amplificar la activación de la AMPK que actúa como una enzima sensible al sustrato que monitorea el nivel de energía celular para impedir consecuencias peores de disminuciones más grandes en el estado de energía [29]-[31]. Este mecanismo dual (disminuciones iniciales en la proporción de ATP/ADP seguida por las disminuciones más tarde en la proporción de ATP/AMP y de ATP/IMP) permitiría que la AMPK advierta el déficit de energía progresivamente sobre un amplio rango de disponibilidad de energía [28].
Las diferencias descritas relacionadas con las repeticiones en la respuesta metabólica aguda a las series repetidas del ejercicio del press de piernas pueden reflejar dos estímulos diferentes para las adaptaciones inducidas por el entrenamiento que ocurren después del entrenamiento pesado de fuerza. Así, algunos estudios han demostrado que el entrenamiento de la fuerza de alta intensidad sin llegar hasta el fallo de los músculos extensores de la rodilla, permite un ambiente favorable para lograr mayores mejoras en la fuerza máxima y la potencia comparado con entrenar hasta el fallo [12], [32], [33]. Tomado en conjunto, los resultados de estos estudios y el presente, indican que un programa de ejercicio de fuerza de extensión de rodillas dinámico caracterizó por la acumulación de bajos metabolitos y mantenimiento de la homeostasis celular y el balance de energía, puede ser una opción más efectiva, eficiente y segura comparado con entrenarse hasta el fallo diseñado para aumentar al máximo la acumulación de fatiga/metabolitos. Bajo esta suposición, sería momento de reemplazar la fiolosofía clásica del entrenamiento "sin dolor, no hay ganancia" a una más racional y lógicamente basada en "sin dolor, más ganancia".
Relaciones entre los metabolitos musculares
Estudios previos han demostrado que durante el ejercicio de extensión de rodillas isométrica y el ejercicio de pedaleo, exhaustivo y no exhaustivo, el contenido absoluto de la PCr cambiaba curvilíneamente y negativamente con respecto al contenido de lactato muscular absoluto [34]. En el presente estudio, una relación negativa curvilínea significativa se observó a lo largo de los ejercicios entre la disminución del porcentaje de la PCr del músculo y el contenido del lactato muscular correspondiente. Esto está de acuerdo con los resultados de Karlsson y colaboradores [35] durante el ejercicio de pedaleo máximo y submáximo y con una fuerte relación entre el logaritmo de la proporción masa-acción de la reacción de la creatina kinasa y el pH muscular reportado en hombres después del ejercicio de extensión de rodillas isométrico [36]. Es más, indica que los cambios entre la PCr y el lactato ocurren principalmente en forma simultánea durante el ejercicio, apoyando las observaciones de otros de que la glucólisis anaeróbica comienza en el músculo al inicio del ejercicio pesado [37]. En acuerdo con nuestros resultados, la acumulación de IMP emparejada cuantitativamente por una disminución en el ATP intramuscular se ha reportado durante el ejercicio de pedaleo submáximo y máximo cuando los niveles de la PCr se caen al 40% de los niveles de reposo [13], [35], [38], [39].
¿Qué causa la fatiga durante las series consecutivas de ejercicio de press de piernas?
Varios autores han indicado que la capacidad para regenerar el ATP a tasas requeridas y así la menor producción de fuerza y de potencia durante el ejercicio máximo de corta duración puede relacionarse a una incapacidad para mantener la tasa requerida de producción anaeróbica de ATP a partir de la degradación de la PCr y del glucógeno, principalmente en las fibras de tipo II [40], [41], un aumento correspondiente en el fosfato inorgánico (Pi) y su forma diprotonada, H2PO4-, aumentos en [H+], alteraciones en el transporte de Ca2+ [42], flujo de K+ desde el músculo [43] o transmisión neuromuscular anormal o fallo de la excitación de membrana [44]. En el presente estudio, la caída en la potencia durante ambos ejercicios en conjunto se correlacionó fuertemente a la caída en los depósitos del ATP y en los niveles de lactato en homogenatos mixtos musculares. La asociación entre los cambios en la potencia y los cambios en la acumulación del lactato muscular está conforme con los estudios antedichos. La asociación observada entre la pérdida de las reservas del ATP y la disminución relativa en la potencia, apoya la idea de que el vaciamiento de ATP en un porcentaje pequeño de fibras puede llevar a su fallo para la producción de potencia [41]. Esta idea coincide con algunos estudios que reportan niveles de ATP post-ejercicio en fibras individuales tan bajos como 1 a 2.4 mmol·Kg-1 de peso muscular húmedo después de un ejercicio de extensión de rodillas máxima [22], o de pedaleo isocinético máximo [41]. De allí, no es irrazonable indicar que algunos cambios bioquímicos, como una disminución en las reservas de ATP y aumentos en el lactato y derivados del ATP (H+, Pi, ADP) de fibras musculares individuales pueden contribuir a la fatiga durante las series sucesivas de ejercicio del press de piernas.
Limitaciones del estudio
Este estudio tuvo algunas limitaciones. Primero, se caracterizó por un número bajo de sujetos experimentales. Sin embargo, las fuertes diferencias estadísticamente significativas observadas en los metabolitos musculares y la potencia entre los dos ejercicios indican que es poco probable que estas diferencias ocurrieran por casualidad. Los estudios longitudinales de biopsia futuros deben buscar reclutar un número mayor de sujetos experimentales para reducir el riesgo potencial errores de tipo II. Segundo, también puede defenderse que el arreglo experimental podría fortalecerse coincidiendo la misma carga a lo largo de las series y el período de descanso total entre los protocolos (por ejemplo, la pausa de 1 minuto entre las series en 5REP y la pausa de 2 minutos entre las series en 10REP). Esto habilitaría comparaciones entre dos tipos de ejercicio de igual volumen. Es obvio que, en este caso, la magnitud de las diferencias entre los ejercicios podría ser inferior que la que se observó en el presente estudio. Sin embargo, tiene que ser recalcado que la meta principal era comparar los cambios metabólicos y de la potencia durante dos tipos de ejercicios de fuerza tradicionalmente usados para alcanzar resultados de entrenamiento específicos ("hipertrofia" vs "mejora óptima de fuerza"), pero no comparar dos ejercicios de igual volumen. Finalmente, este estudio se realizó en hombres entrenados en resistencia con una proporción alta (65%) de fibras de tipo I. Debe tenerse cuidado al generalizar los resultados de este estudio a otras poblaciones (por ejemplo, atletas de potencia con una proporción alta de fibras de tipo II). A pesar de estas limitaciones, los resultados proveen importante y nueva información sobre las características metabólicas de dos ejercicios de fuerza de alta intensidad muy populares diseñados para aumentar la fuerza muscular, la potencia e hipertrofia en atletas y adultos con enfermedades crónicas e invalideces. Esto habilitaría comparaciones entre dos tipos de ejercicio de igual volumen. Es obvio que, en este caso, la magnitud de las diferencias entre los ejercicios podría ser inferior que la que se observó en el presente estudio. Sin embargo, tiene que ser recalcado que la meta principal era comparar los cambios metabólicos y de la potencia durante dos tipos de ejercicios de fuerza tradicionalmente usados para alcanzar resultados de entrenamiento específicos ("hipertrofia" vs "mejora óptima de fuerza"), pero no comparar dos ejercicios de igual volumen. Finalmente, este estudio se realizó en hombres entrenados en resistencia con una proporción alta (65%) de fibras de tipo I. Debe tenerse cuidado al generalizar los resultados de este estudio a otras poblaciones (por ejemplo, atletas de potencia con una proporción alta de fibras de tipo II). A pesar de estas limitaciones, los resultados proveen importante y nueva información sobre las características metabólicas de dos ejercicios de fuerza de alta intensidad muy populares diseñados para aumentar la fuerza muscular, la potencia e hipertrofia en atletas y adultos con enfermedades crónicas e invalideces.
En resumen, los resultados principales de este estudio fueron, primero, que reduciendo el número de repeticiones durante las series de diez a cinco, manteniendo la misma carga inicial y períodos de recuperación entre las series, indujeron menores demandas en forma notable en el sistema de los fosfatos altos en energía y en el aporte de energía glucolítica, síntesis del ATP, que permitió coincidir con la utilización del ATP, mantener la potencia y el balance de energía a lo largo de las series y experimentar mucho menos fatiga y disconfort durante el ejercicio del press de piernas bilateral. Segundo, en los homogenatos mixtos del músculo se correlacionaron los cambios en la PCr del músculo durante ambos ejercicios con el lactato muscular y el contenido de IMP. Estas correlaciones indican que los cambios entre la PCr y el lactato muscular ocurren principalmente en forma simultánea visto que el IMP solamente se acumula cuando los niveles de PCr son bajos. Finalmente, la caída en la producción de la potencia se correlacionó fuertemente a la caída en las reservas de ATP y a los niveles del lactato muscular. Esto indica que una disminución en la reserva de ATP y aumentos en el lactato y de los derivados de ATP (H+, Pi, ADP) de fibras musculares individuales pueden contribuir a la fatiga durante series sucesivas de ejercicio del press de piernas.
Cita: Gorostiaga EM, Navarro-Amézqueta I, Calbet JAL, Hellsten Y, Cusso R, Guerrero M, et al. (2012) Energy Metabolism during Repeated Sets of Leg Press Exercise Leading to Failure or Not. PLoS ONE 7(7): e40621.
Editor: Nuria Garatachea, University of Zaragoza, Spain
Recibido: Febrero 26, 2012; Aceptado: Junio 11, 2012; Publicado: Julio 13, 2012
Copyright: © 2012 Gorostiaga et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This study was supported by the Spanish Ministry of Education (National Plan of R&D+I 2004–2007, Key action “Sport and Physical Activity” DEP2006-56076). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
REFERENCIAS
1. Colberg SR, Sigal RJ, Fernhall B, Regensteiner JG, Blissmer BJ, et al. (2010). Exercise and type 2 diabetes: the American College of Sports Medicine and the American Diabetes Association: joint position statement. Diabetes Care 33: e147–e167.
2. Kraemer WJ, Adams K, Cafarelli E, Dudley GA, Dooly C, et al. (2002). American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. Med Sci Sports Exerc 34: 364–380.
3. Hancock CR, Janssen E, Terjung RL (2006). Contraction-mediated phosphorylation of AMPK is lower in skeletal muscle of adenylate kinase-deficient mice. J Appl Physiol 100: 406–413.
4. Winder WW, Hardie DG (1999). AMP-activated protein kinase, a metabolic master switch: possible roles in type 2 diabetes. Am J Physiol 277: E1–10.
5. McCartney N, Spriet LL, Heigenhauser GJ, Kowalchuk JM, Sutton JR, et al. (1986). Muscle power and metabolism in maximal intermittent exercise. J Appl Physiol 60: 1164–1169.
6. Saltin B, Essén B (1971). Muscle glycongen, lactate, ATP, and CP in intermittent exercise. In: Pernow B, Saltin B, editors. pp. 419–424. New York: Plenum.
7. Cheetham ME, Boobis LH, Brooks S, Williams C (1986). Human muscle metabolism during sprint running. J Appl Physiol 61: 54–60.
8. Edwards RH, Harris RC, Hultman E, Kaijser L, Koh D, et al. (1972). Effect of temperature on muscle energy metabolism and endurance during successive isometric contractions, sustained to fatigue, of the quadriceps muscle in man. J Physiol 220: 335–352.
9. Spriet LL, Lindinger MI, McKelvie RS, Heigenhauser GJ, Jones NL (1989). Muscle glycogenolysis and H+ concentration during maximal intermittent cycling. J Appl Physiol 66: 8–13.
10. Burd NA, Andrews RJ, West DW, Little JP, Cochran AJ, et al. (2012). Muscle time under tension during resistance exercise stimulates differential muscle protein sub-fractional synthetic responses in men. J Physiol 590: 351–362.
11. Kraemer WJ, Nobel BJ, Clarck MJ, Culver BW (1987). Physiological responses to heavy-resistance exercise with very short rest period. Int J Sports Med 8: 247–252.
12. Izquierdo M, Ibanez J, Gonzalez-Badillo JJ, Hakkinen K, Ratamess NA, et al. (2006). Differential effects of strength training leading to failure versus not to failure on hormonal responses, strength, and muscle power gains. J Appl Physiol 100: 1647–1656.
13. Sahlin K, Broberg S (1990). Adenine nucleotide depletion in human muscle during exercise: causality and significance of AMP deamination. Int J Sports Med 11: S62–S67.
14. Izquierdo M, Gonzalez-Badillo JJ, Hakkinen K, Ibanez J, Kraemer WJ, et al. (2006). Effect of loading on unintentional lifting velocity declines during single sets of repetitions to failure during upper and lower extremity muscle actions. Int J Sports Med 27: 718–724.
15. Storer TW, Davis JA, Caiozzo VJ (1990). Accurate prediction of VO2max in cycle ergometry. Med Sci Sports Exerc 22: 704–712.
16. Bergstrom J (1975). Percutaneous needle biopsy of skeletal muscle in physiological and clinical research. Scand J Clin Lab Invest 35: 609–616.
17. Lowry OH, Passonneau JV (1971). Some recent refinements of quantitative histochemical analysis. Curr Probl Clin Biochem 3: 63–84.
18. Norman B, Heden P, Jansson E (1991). Small accumulation of inosine monophosphate (IMP). despite high lactate levels in latissimus dorsi during transplantation. Clin Physiol 11: 375–384.
19. Brooke MH, Kaiser KK (1969). Some comments on the histochemical characterization of muscle adenosine triphosphatase. J Histochem Cytochem 17: 431–432.
20. Hellsten Y, Sjodin B, Richter EA, Bangsbo J (1998). Urate uptake and lowered ATP levels in human muscle after high-intensity intermittent exercise. Am J Physiol 274: E600–E606.
21. Tesch PA, Colliander EB, Kaiser P (1986). Muscle metabolism during intense, heavy-resistance exercise. Eur J Appl Physiol Occup Physiol 55: 362–366.
22. Jansson E, Dudley GA, Norman B, Tesch PA (1987). ATP and IMP in single human muscle fibres after high intensity exercise. Clin Physiol 7: 337–345.
23. Casey A, Constantin-Teodosiu D, Howell S, Hultman E, Greenhaff PL (1996). Metabolic response of type I and II muscle fibers during repeated bouts of maximal exercise in humans. Am J Physiol 271: E38–E43.
24. Koopman R, Manders RJ, Jonkers RA, Hul GB, Kuipers H, et al. (2006). Intramyocellular lipid and glycogen content are reduced following resistance exercise in untrained healthy males. Eur J Appl Physiol 96: 525–534.
25. Bogdanis GC, Nevill ME, Boobis LH, Lakomy HK (1996). Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J Appl Physiol 80: 876–884.
26. Parolin ML, Chesley A, Matsos MP, Spriet LL, Jones NL, et al. (1999). Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am J Physiol 277: E890–E900.
27. Howlett RA, Parolin ML, Dyck DJ, Hultman E, Jones NL, et al. (1998). Regulation of skeletal muscle glycogen phosphorylase and PDH at varying exercise power outputs. Am J Physiol 275: R418–R425.
28. Xiao B, Sanders MJ, Underwood E, Heath R, Mayer FV, et al. (2011). Structure of mammalian AMPK and its regulation by ADP. Nature 472: 230–233.
29. Hancock CR, Brault JJ, Terjung RL (2006). Protecting the cellular energy state during contractions: role of AMP deaminase. J Physiol Pharmacol 57: 17–29.
30. Hardie DG, Carling D (1997). The AMP-activated protein kinase–fuel gauge of the mammalian cell? Eur J Biochem 246: 259–273.
31. Tullson PC, Terjung RL (1991). Adenine nucleotide metabolism in contracting skeletal muscle. Exerc Sport Sci Rev 19: 507–537.
32. Kraemer WJ, Stone MH, O’Bryant HS, Conley MS, Johnson RL, et al. (1997). Effects of single vs. multiple sets of weight training: Impact of volume, intensity, and variation. J Strength Cond Res 11: 143–147.
33. Sanborn K, Boros R, Hruby J, Schilling B, O’Bryant HS, et al. (2000). Short-term performance effects of weight training with multiple sets not to failure vs. a single set to failure in women. J Strength Cond Res 14: 328–331.
34. Harris RC, Sahlin K, Hultman E (1977). Phosphagen and lactate contents of m. quadriceps femoris of man after exercise. J Appl Physiol 43: 852–857.
35. Karlsson J, Diamant B, Saltin B (1970). Muscle metabolites during submaximal and maximal exercise in man. Scand J Clin Lab Invest 26: 385–394.
36. Sahlin K, Harris RC, Hultman E (1975). Creatine kinase equilibrium and lactate content compared with muscle pH in tissue samples obtained after isometric exercise. Biochem J 152: 173–180.
37. Gollnick PD, Hermansen L (1973). Biochemical adaptations to exercise: anaerobic metabolism. Exerc Sport Sci Rev 1: 1–43.
38. Hultman E (1967). Studies on muscle metabolism of glycogen and active phosphate in man with special reference to exercise and diet. Scand J Clin Lab Invest. pp. 1–63.
39. Spriet LL, Soderlund K, Bergstrom M, Hultman E (1987). Anaerobic energy release in skeletal muscle during electrical stimulation in men. J Appl Physiol 62: 611–615.
40. Hultman E, Greenhaff PL (1991). Skeletal muscle energy metabolism and fatigue during intense exercise in man. Sci Prog 75: 361–370.
41. Karatzaferi C, de HA, Ferguson RA, van MW, Sargeant AJ (2001). Phosphocreatine and ATP content in human single muscle fibres before and after maximum dynamic exercise. Pflugers Arch 442: 467–474.
42. Nakamaru Y, Schwartz A (1972). The influence of hydrogen ion concentration on calcium binding and release by skeletal muscle sarcoplasmic reticulum. J Gen Physiol 59: 22–32.
43. Juel C (1988). Muscle action potential propagation velocity changes during activity. Muscle Nerve 11: 714–719.
44. Edwards RHT (1981). Human muscle function and fatigue. In: Human Muscle Fatigue: Physiological mechanisms. London: Pitman. pp. 1–18.
Evaluación de la integridad de los datos de registros médicos electrónicos en tiempo real utilizados en la investigación clínica.
Resumen Antecedentes Los datos de registros médicos electrónicos (EHR) casi en tiempo real ofrecen...

La influencia de la configuración espacial del jardín en el comportamiento turístico: una revisión sistemática basada en la sintaxis espacial
ResumenComo espacios compuestos que integran naturaleza y cultura, los jardines ya no se...

Factores domésticos que influyen en las infestaciones de cucarachas y parásitos helmintos: perspectivas de una comunidad rural en Guatemala
ResumenLas cucarachas son vectores de patógenos y parásitos que plantean riesgos para la salud...
Corrección: un método analítico GC-MS-SIM mejorado para la determinación de residuos de pendimetalina en cultivos comerciales (hojas y suelos) y su validación
Hay errores en la declaración de financiación. La declaración de financiación correcta es la...
Más allá de la percepción de la desigualdad económica actual: cómo interpretan las personas las brechas de riqueza pasadas, presentes y futuras
ResumenLa gente tiende a percibir erróneamente los niveles de desigualdad económica en su país....