Resumen
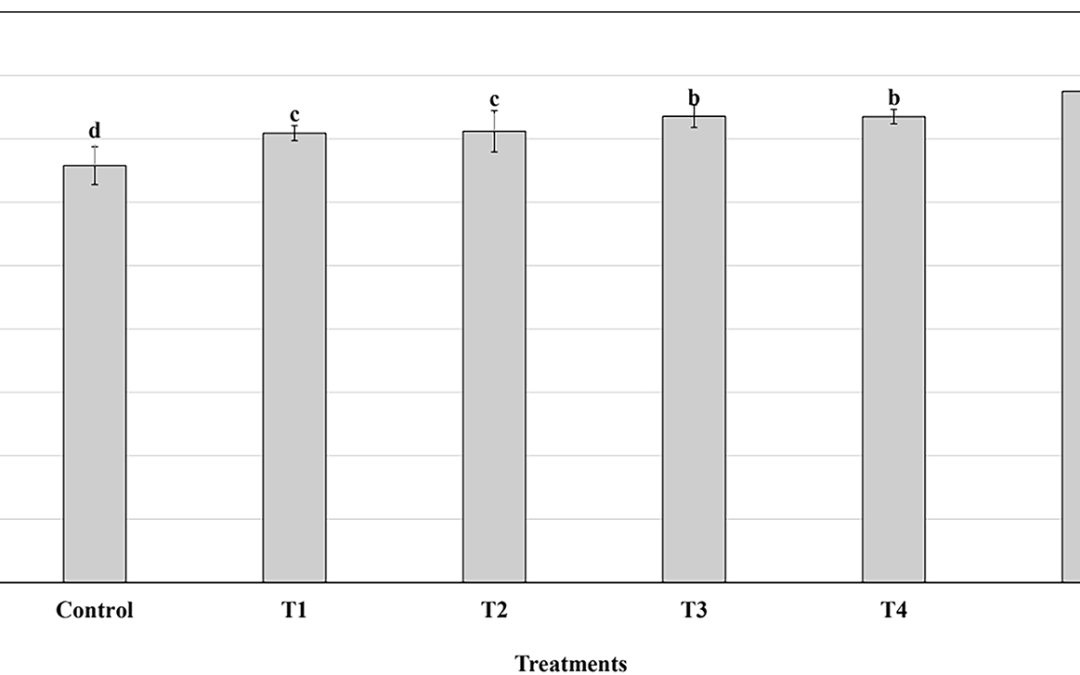
El entrenamiento de fuerza unilateral puede causar efectos de fuerza de transferencia cruzada a los músculos contralaterales homólogos. Sin embargo, el impacto de los efectos cruzados en el tejido muscular no está claro. Para probar la hipótesis de que el uso excesivo unilateral del músculo causa alteraciones bilaterales en la composición de la fibra muscular y el suministro vascular, hemos utilizado un modelo de conejo experimental con ejercicio unilateral de exceso de estraz a través de la estimulación muscular eléctrica (E/EMS). Los músculos Soleus (SOL) y Gastrocnemius (GA) de las piernas ejercidas (E) y contralaterales no ejercidas (NE) (n = 24) se analizaron morfológicamente después de 1W, 3W y 6W de EMS. Los conejos no ejercidos sirvieron como controles (n = 6). Después de la intervención unilateral, los músculos de las piernas E y NE mostraron miositis y cambios de tejido estructural y molecular que a varios grados se reflejaban entre sí. El área de fibra era bilateralmente más pequeña que en los controles después de 3W de E/EMS en SOL (E 4420 y NE 4333 µm2 vs. 5183 µm2p <0.05) y GA (E 3572 y NE 2983 µm2 vs. 4697 µm2p <0.02) músculos. Después de 6W de E/EMS, el porcentaje de fibras MYHCI lentas fue menor que en los controles en las patas NE de SOL (88.1% frente a 98.1%, p <0.009), mientras que el porcentaje de fibras de Myhciia rápidas fue mayor en las piernas NE de GA (25.7% vs. 15.8%, P = 0.02). El número de capilares alrededor de las fibras en las piernas E y NE fue menor (SOL 13% y 15%, respectivamente, GA 25% y 23%, respectivamente, P <0.05) que en los controles. Las alteraciones generales fueron más marcadas en el músculo de GA rápido que en el músculo SOL lento, que por otro lado mostró más cambios musculares histopatológicos. Llegamos a la conclusión de que el ejercicio de uso excesivo de uso exagerado unilateral descargado a través de EMS causa miositis y cambios musculares en las proporciones de tipo de fibra, el área de fibra y la capilarización de la fibra no solo en la pierna ejercida, sino también en los músculos homólogos de la pierna no ejercida.
Introducción
Es ampliamente aceptado que el entrenamiento de fuerza unilateral aumenta la fuerza no solo en el músculo entrenado sino también en cierta medida en el músculo de los homólogos de la extremidad contralateral (1)–(4). Este efecto se ha informado tanto para los músculos de la extremidad pequeños y grandes y puede ocurrir por diversas modalidades del ejercicio (e) que se realiza por los esfuerzos voluntarios, así como por la estimulación muscular eléctrica (EMS). (5)–(10). El potencial del efecto para inducir la mejora de la fuerza en el músculo contralateral no entrenado es de gran interés y tiene una relevancia obvia en la rehabilitación clínica. Por ejemplo, se ha informado que el ejercicio unilateral previene las extremidades inmovilizadas contralaterales en individuos sanos por pérdida de masa muscular y fuerza (11). Incluso un episodio de ejercicio excéntrico puede tener un efecto protector en la extremidad contralateral (12), (13). También se ha demostrado que el ejercicio inducido a través de EMS dio como resultado mayores efectos contralaterales que el inducido por la capacitación de contracción voluntaria (4), (9), (14). Sin embargo, los eventos adversos también pueden transferirse cruzados. Recientemente informamos que el musgo unilateral en exceso por E/EMS que conduce a una lesión en el músculo de los tríceps de conejo también afectó el músculo contralateral homólogo. En ese estudio encontramos cambios musculares histopatológicos focales e inflamación (miositis) tanto en la pierna manipulada como en la de reposo (15). Además, hubo una regulación positiva bilateral de la sustancia neuropéptido P, un neuromodulador en la familia de taquiquinina que se sabe que está involucrado en la inflamación neurogénica y la vasodilatación (16). Se han informado cambios contralaterales en el sistema nervioso periférico después de una lesión nerviosa unilateral en numerosos estudios (para revisión ver (17)), los hallazgos que respaldan la idea de que los eventos nocivos en un músculo pueden ser transferidos. El apoyo adicional para una actividad de transferencia cruzada del sistema nervioso es la distribución a menudo estrictamente simétrica de la artritis reumatoide y algunas enfermedades inflamatorias crónicas y una imagen espejo del sistema nervioso para el dolor. (18)–(21). Aunque los efectos cruzados están bien descritos en la literatura, no está claro si el ejercicio de uso excesivo unilateral causa cambios estructurales o moleculares en los músculos contralaterales.
Los músculos esqueléticos tienen un potencial adaptativo para modificar su composición de los fenotipos musculares y su perfil metabólico en respuesta a patrones de actividad alterados (22)–(24). Esta respuesta adaptativa refleja, entre varios otros factores, la capacidad de una célula muscular para modular el tipo de isoformas de cadena pesada de miosina contráctil (MYHC), tamaño de las fibras (25)capacidad oxidativa mitocondrial y suministro capilar de las células musculares (26)–(29). Hasta donde sabemos, ningún estudio ha examinado sistemáticamente las alteraciones en la expresión de las principales proteínas contráctiles MyHC, el tamaño de la fibra muscular y el suministro capilar cuando los músculos contralaterales han sido influenciados por el ejercicio a través de EMS.
En el presente estudio, se usó un modelo de ejercicio de conejo para probar la hipótesis de que el exceso de exceso descargado unilateral inducido por E/EMS no solo causa alteraciones bilaterales de tejido histopatológico e inflamación (15), (16)pero también cambios bilaterales en el sistema motor contráctil y en el compartimento vascular. El músculo tríceps surae de una pierna estuvo expuesto a E/EMS repetidos durante tres períodos experimentales, 1, 3 y 6 semanas. Después de cada período experimental, los músculos ejercidos y no ejercidos se analizaron con enzimas y técnicas inmunohistoquímicas y morfológicas para determinar posibles cambios en las composiciones de fenotipo de fibra, la morfología de la fibra y el suministro capilar. La ventaja de usar el músculo tríceps surae es que sus dos partes musculares principales, los músculos sóleo y gastrocnemio, en varias especies difieren sustancialmente entre sí en la composición del tipo de fibra muscular (30), (31).
Materiales y métodos
Declaración de ética
El estudio fue aprobado por el comité ético de la Universidad de Umeå (Protocolo A34/07) que cumplió con las directrices y estándares nacionales (SFS1988: 534; 1988: 539) e International (2010/63/UE) en la investigación animal. La aprobación se obtuvo antes del inicio del estudio. Un criador con licencia había criado a todos los animales con el único propósito de ser utilizado en experimentos con animales. Se hicieron todos los esfuerzos para minimizar el sufrimiento animal.
Animales
El experimento se llevó a cabo en conejos blancos de Nueva Zelanda de 6 a 9 meses y pesaba aproximadamente 4 kg. En total, se incluyeron 24 conejos en el experimento, de los cuales 18 se distribuyeron en tres grupos experimentales y los otros sirvieron como controles (6 conejos en cada grupo). A lo largo del experimento, los conejos se mantuvieron bajo anestesia, inducidos por inyecciones intramusculares de diazepham (0.2 ml/kg) y fentanilfluanisona (0.2-0.3 ml/kg). Para mantener la anestesia, se inyectó más fentanilfluanison (0.1 ml/kg) cada 30-45 minutos durante el procedimiento del experimento. La buprenorfina (0.01–0.05 mg/kg) recibió SC después de la operación para minimizar el dolor. Los seis animales que constituyen el grupo de control se mantuvieron en jaulas ordinarias, pero no habían sido sometidos al régimen de ejercicio en el experimento.
Diseño experimental
Se usó un modelo experimental que usa una máquina de patada, construido para el ejercicio descargado a través de la estimulación muscular eléctrica simultánea de tres conejos, para el logro de flexiones pasivas y extensiones de la articulación del tobillo derecho. Este modelo es una forma modificada del modelo utilizado por Backman et al. (32), (33)y eso se ha utilizado previamente para examinar los cambios en el tendón (tendinopatía/tendinosis) en el tendón de Aquiles (32), (33). Un pistón neumático unido al pie derecho produjo los movimientos de la articulación del tobillo, en el que se podía controlar el rango de movimiento. El rango de movimiento se estableció en 9.5 cm, dado un rango de movimiento en el tobillo de 55-65 °, de los cuales 20–25 ° era dorsiflexión y 35-40 ° era flexión plantar. Una banda estaba atada alrededor de la pelvis/cadera del conejo para restringir el movimiento en la pierna no ejercitada izquierda. Durante la flexión plantar, una contracción concéntrica activa fue inducida por la estimulación eléctrica a través de electrodos superficiales (electrodo pediátrico 40 426a, Helwett Packard, Andover, MA, EE. UU.), Se colocaron 2 cm de distancia sobre el músculo tríceps surae. La estimulación se sincronizó con el movimiento de flexión plantar del pistón mediante un microeswitch, que trigó la unidad de estimulador (Disa estimulador tipo 14e 10; disa elektronik A/S, Herlev, Dinamarca). El estimulador generó un impulso con una duración de 0.2 ms que se administró 85 ms después del inicio de la flexión plantar a una amplitud de 35 a 50 V. El estímulo se controló por un oscilómetro y la frecuencia de la flexión y los movimientos de extensión se estableció en 150 por minuto (2.5 Hz) y el movimiento repetitivo se estableció en un período de 2 horas. El experimento se repitió cada segundo día y la duración de los períodos experimentales fue de 1 semana, 3 semanas y 6 semanas. Los conejos se mantuvieron en jaulas ordinarias que permitían la libertad de movimiento entre los períodos experimentales. Después de cada período experimental, los animales fueron sacrificados (el día después de la última sesión de experimento) por una sobredosis intravenosa de pentobarbital. Por lo tanto, los animales examinados después de un período experimental de 1 semana habían sido sometidos a las sesiones experimentales en cuatro ocasiones. Los animales de control fueron sacrificados en relación con el inicio de los experimentos. Para más detalles, ver Backman et al. (32)Andersson et al. (33) y Song et al. (15).
Muestras musculares
Después de que los animales fueron sacrificados, el músculo tríceps surae se disecó de los lados experimentales y contralaterales. Se identificaron los músculos Soleus y Gastrocnemius de los tríceps surae y se obtuvieron muestras musculares de un tamaño aproximado de 5 × 5–10 mm de la misma área en las porciones distales de los músculos de ambas piernas. En el músculo gastrocnemio, la muestra se obtuvo de la parte central/distal del músculo, mientras que la muestra del músculo sóleo representaba toda la sección transversal del vientre muscular. Las muestras musculares se montaron directamente en el compuesto OCT (tejido TEK, Miles Laboratories, Naperville, IL, EE. UU.) En un cartón delgado y se congelaron rápidamente en propano enfriado con nitrógeno líquido, y se almacenaron a -80 ° hasta su uso. Se cortaron secciones transversales seriales de cinco y 8 µm de espesor en un microtomo de criostato (RecHert-jung, Leica Heidelberg, Alemania) a -23 ° C y se montaron en portaobjetos de vidrio.
Inmunohistoquímica
Las secciones transversales musculares de 5 µm de grosor se procesaron para inmunhistochemistry (IHC) con anticuerpos monoclonales (MAB) previamente caracterizados y un anticuerpo policlonal (AB). Las principales proteínas contráctiles en las fibras musculares, las isoformas de la cadena pesada de miosina (MYHC), se analizaron utilizando diferentes mAbs contra isoformas de MYHC de desarrollo y adulto (para detalles de anticuerpos y origen ver Tabla 1) (34)–(37). La visualización del contorno de la fibra muscular (es decir, la lámina basal) de las fibras musculares se realizó con tinción para la PC128 policlonal contra la laminina (grupo de unión, el Reino Unido, diluido 1∶15000) y MAb NCL-merosin contra laminina α2 α2 (Novocastra Laboratories Ltd, UK, diluido 1∶1000)) (38). Se usó MAB D33 (Dako, Dinamarca, diluido 1∶1000) contra la desmin para la detección de degeneración de la fibra muscular (aumento de la tinción) y la necrosis de fibra (sin tinción) y MAB 1891 contra la fibronectina (Chemicon, Temecula, CA, EE. UU., Se utilizaron 1∶2000 diluidos) para el análisis de la fibrosis. Para identificar capilares, MAB M0823 contra CD31 (Dako Glostrup, Dinamarca, diluido 1∶100) que reconoce PECAM-1 (molécula de adhesión endotelial de plaquetas), se utilizó una glucoproteína transmembranosa fuertemente expresada por células endoteliales vasculares. Se ha demostrado previamente que los mAbs contra CD31 identifican el endotelio vascular en el músculo normal (39), (40) y en tendones de conejo (33). Los axones en los nervios se tiñeron con MAB T8660 contra la tubulina β-III (Sigma-Aldrich, EE. UU., Diluido 1∶500) y las células Schwann se tiñeron con MAB S2532 contra S-100beta (Sigma-Aldrich, EE. UU., Diluido 1∶500) (15), (41), (42).