
Pew Center for Molecular Nutrition, and Department of Nutrition, Case Western Reserve University School of Medicine Cleveland, OH 44106.
Resumen
ROL DEL SUMINISTRO DE SUSTRATOS
Luego del ejercicio prolongado, las reservas de glucógeno muscular permanecen bajas hasta que la alimentación con carbohidratos es iniciada. La resíntesis de glucógeno es más rápida si los carbohidratos son administrados las dos primeras horas después del ejercicio. Los investigadores que examinaron la cantidad de carbohidratos consumida después del ejercicio han encontrado que un incremento en el consumo de carbohidratos hasta 648 g cada 24 horas, resulto en cantidades progresivamente mayores de resíntesis de glucógeno muscular en sujetos en los cuales sus niveles de glucógeno habían sido disminuidos por medio de una carrera de 16.1 km al 80 % del VO2 máx. (1). Otros investigadores han encontrado que existe un límite superior en la síntesis de glucógeno, cuando se consumen carbohidratos. En sujetos que doblaron el consumo de carbohidratos de 0.35 g/ kg de peso corporal hasta 0.7 g/kg de peso corporal, la síntesis de glucógeno aumento más del doble durante las primeras 24 horas después del ejercicio. Sin embargo, si el consumo de carbohidratos era incrementado hasta 1.5 a 3.0 g de glucosa /kg de peso corporal, no era observado un mayor incremento en la velocidad de resíntesis de glucógeno.
Estos estudios sugieren que las grandes cargas de glucosa pueden enlentecer el vaciado y la absorción gástrica, y por esto, limitar la repleción del glucógeno muscular después del ejercicio.
Han sido investigados el tipo de carbohidratos ingeridos después del ejercicio, para determinar si los carbohidratos simples vs. complejos afectan la velocidad de resíntesis de glucógeno. En sujetos a los que se les dio fructosa durante la recuperación, la resíntesis de glucógeno en el músculo fue reducida en comparación con sujetos a los que se les dio glucosa. Sin embargo, en ratas a las que se les dio fructosa en vez de glucosa durante la recuperación, a pesar de que el glucógeno muscular era menor, el nivel de glucógeno alcanzado en el hígado fue tres veces mayor en comparación con las ratas alimentadas a glucosa.
El mayor nivel de actividad de la fructo quinasa en el hígado comparado con el relativamente bajo nivel en el músculo, explica más probablemente la deposición incrementada del glucógeno hepático. En los sujetos que se alimentaron con carbohidratos simples vs. complejos, los niveles de glucógeno muscular fueron más altos durante las primeras 6 horas después del ejercicio, sugiriendo que el proceso de digestión y transporte de glucosa conduce a síntesis de glucógeno más rápidas. En un intento de evitar las limitaciones impuestas por la digestión de la glucosa, los investigadores administraron glucosa intravenosa y compararon la velocidad de repleción de glucógeno post-ejercicio con sujetos a los que se les había dado glucosa oralmente. Cuando los niveles plasmáticos de glucosa fueron doblados durante un protocolo de infusión de glucosa (1g.kg-¹.hr-¹) la velocidad de resíntesis de glucógeno muscular fue 2 a 3 veces más alta que cuando la glucosa fue administrada oralmente. Resultados similares fueron encontrados cuando los niveles de insulina fueron mantenidos constantes por medio de la técnica de atrapamiento de la insulina, combinada con la infusión de glucosa. Así, las investigaciones disponibles sugieren que la velocidad de resíntesis de glucógeno está relacionada a los niveles de glucosa plasmática, mientras que la ruta de administración es menos importante.
ROL DEL HÍGADO EN LA DISPOSICIÓN DE LOS CARBOHIDRATOS
Durante la transición del estado de ejercicio y ayuno a la condición de reestablecimiento del glucógeno, el hígado juega un rol clave para dirigir el suministro de glucosa ingerida al músculo esquelético, para su deposición. Aunque la deposición de glucosa ocurre predominantemente en las células musculares, la extensión a la cual el glucógeno muscular es derivado por medio de la captación y fosforilación de glucosa, y a partir de orígenes distintos a la glucosa, tal como el lactato y piruvato, puede variar dependiendo de las condiciones metabólicas. Es conocido que la glucosa que es absorbida por el intestino, conduce a la recuperación del glucógeno en el músculo mucho más temprano que en el hígado.
Ha sido sugerido, a partir de estudios con infusión de trazadores dentro del lactato en animales, una contribución significativa del ácido láctico a la deposición de glucógeno muscular. En el animal intacto, una proporción del ácido láctico producido durante el ejercicio, es utilizado en los músculos inactivos. Sin embargo, el transporte de lactato y otros sustratos al hígado durante el ejercicio, resulta en una producción acelerada de glucosa hepática, por medio de la ruta metabólica conocida como gluconeogénesis.
La gluconeogénesis en el hígado comprende una serie de pasos enzimáticos inversos a la ruta de la glucólisis. Los sustratos requeridos para este proceso incluyen, lactato, glicerol y al aminoácido alanina. Durante el ejercicio estos sustratos son derivados del metabolismo muscular y adiposo, entran a la sangre, son extraídos por el hígado y reciclados a través del proceso gluconeogénico para producir glucosa. Esta ruta es necesaria para mantener la glucosa sanguínea durante el ejercicio debido a la utilización incrementada de glucosa por parte de los músculos esqueléticos durante el ejercicio. El incremento de la gluconeogénesis durante el ejercicio es debido en parte a la activación de enzimas gluconeogénicas en el hígado, por las hormonas glucagón, epinefrina, y cortisol, y por el transporte incrementado de sustratos. Durante la recuperación, estas hormonas tienden a disminuir, y la insulina, que suprime la gluconeogénesis, se incrementa durante la alimentación. Sin embargo, la contribución del lactato a la gluconeogénesis, puede continuar siendo alta, si los niveles de insulina son mantenidos bajos, y los niveles de sustratos, particularmente el lactato, permanecen elevados. Además, hay evidencia de estudios incubados in-vitro, que los músculos de las fibras rápidas, en particular, son capaces de formar glucosa a partir de ácido láctico, un proceso al que se le atribuye ser la ruta indirecta de resíntesis de glucógeno. In-vivo, la infusión de glucosa marcada con 14C después del ejercicio, produjo un incremento marcado en el lactato-14C, y contribuyo a la formación de glucógeno marcado con 14C, depositado en la porción blanca del gastrocnemio (2). Esto se opone a lo que ocurre en los músculos de fibras lentas, que sintetizan glucógeno casi exclusivamente a partir de la glucosa plasmática por medio de la captación directa y deposición, llamada de otro modo, la ruta directa. En ratas que hacían ayuno, la rápida recuperación de la glucosa sanguínea después de la hipoglucemia producida por el ejercicio, sugirió que el lactato puede también comportarse como un origen de glucosa, en vez de ser usado como un sustrato para la oxidación durante la recuperación.
EVENTOS BIOQUÍMICOS IMPLICADOS EN LA DEPOSICIÓN DE GLUCÓGENO – EL ROL DE LA GLUCÓGENO SINTETASA
Las velocidades de deposición de glucógeno después del ejercicio parecen ser directamente proporcionales a la extensión a la cual ha ocurrido la depleción de glucógeno durante el ejercicio.
Además, el patrón de repleción de glucógeno después del ejercicio es más rápido y más alto en los músculos compuestos principalmente por tipos de fibras oxidativas. Esto ha sugerido ser un reflejo de la mayor actividad de las enzimas hexoquinasa y glucógeno sintetiza en las fibras rojas rápidas y lentas, que en las fibras blancas rápidas.
El ejercicio y la insulina inducen por si mismos, un incremento en la actividad de la enzima glucógeno sintetasa, por medio de un incremento en el porcentaje de enzima en la forma I (independiente), lo que significa que la enzima puede ser activada independientemente de la glucosa-6-fosfato, el principal modificador alostérico de la glucógeno sintetasa. Cuando desciende la concentración de glucógeno en el músculo, la enzima glucógeno sintetasa es liberada del glucógeno y está disponible para ser desfosforilada por la enzima fosfoproteín fosfatasa, que cataliza la conversión de la sintetasa D (forma dependiente) a la forma I, forma activa más espontánea.
Aunque pequeños cambios en la proporción de actividad, de D a I, pueden conducir a grandes cambios en la velocidad de síntesis del glucógeno, la glucógeno sintetiza I, parece incrementarse muy poco en respuesta a la depleción de glucógeno. Algunos estudios han sugerido que la resíntesis de glucógeno puede ser el resultado de una pequeña, pero significativa activación de la glucógeno sintetasa en la forma D, en tales condiciones, bajos niveles de glucosa-6-fosfato son necesarios para estimular la actividad de tal enzima. La acumulación de glucosa-6-fosfato en el músculo esquelético esta limitada en parte por la velocidad en que la glucosa entra en la célula muscular. Aunque la velocidad acelerada de la resíntesis de glucógeno durante la recuperación inicial después del ejercicio comprende la activación de la enzima glucógeno sintetasa, el ejercicio parece estimular la síntesis de glucógeno muscular más de 8 veces luego del ejercicio intenso, sin embargo la actividad de la glucógeno sintetasa se incrementa solo aproximadamente un 25 %. Estas observaciones indican que la formación de glucógeno no depende solamente de grandes incrementos de la actividad de la glucógeno sintetasa, y sugiere que los factores que incrementan la captación de glucosa en la célula muscular, pueden ser más críticos para la resíntesis de glucógeno después del ejercicio.
ROL DEL TRANSPORTE DE GLUCOSA EN LA REGULACIÓN DE LA SÍNTESIS DE GLUCÓGENO
Bajo la mayoría de las condiciones metabólicas, el paso limitante en el metabolismo de la glucosa muscular, parece ser el transporte de la glucosa. Así, los factores que incrementan el transporte de la glucosa dentro del músculo esquelético, tienen probablemente un efecto significativo sobre la síntesis del glucógeno muscular. El ejercicio o la actividad contráctil, son un potente estímulo para incrementar la permeabilidad del músculo a la glucosa, aún en la ausencia de insulina. Los reportes indican que la captación de glucosa puede incrementarse tanto como 10 veces en la ausencia de insulina, luego de una hora de ejercicio de intensidad moderada. Tiene mérito señalar que el transporte incrementado de glucosa luego de contracciones musculares, ocurre solo en músculos que están significativamente depletados de glucógeno y que el transporte de glucosa retorna a niveles normales en músculos que han sido repletados de glucógeno. El trasporte incrementado de glucosa dentro del músculo persiste por 16 a 20 horas luego del ejercicio en ratas, si es restringida la ingesta de carbohidratos. Sin embargo, la velocidad de transporte de glucosa no es incrementada cuando el glucógeno muscular es disminuido por medio de una noche de ayuno. Esto sugiere que la actividad contráctil provee un estímulo importante para conectar el transporte de glucosa a la síntesis incrementada de glucógeno muscular después del ejercicio. Aunque la naturaleza exacta de la señal biológica requerida para el incremento en el transporte de glucosa dentro del músculo, inducido por el ejercicio, permanece indefinida, el sistema efector que facilita la entrada de glucosa dentro del músculo ha sido recientemente identificado.
ROL DE LOS TRANSPORTADORES DE GLUCOSA EN LA CAPTACIÓN DE GLUCOSA DESPUÉS DEL EJERCICIO
Mientras que los estudios clásicos de fisiología y bioquímica, han enfatizado la importancia de la actividad de la enzima glucógeno sintetasa, para explicar al resíntesis de glucógeno muscular después del ejercicio, el conocimiento de las bases moleculares de la captación de glucosa en los músculos se ha expandido enormemente, luego de la clonación y caracterización de la familia de las proteínas transportadoras de glucosa. Las mismas pueden ser detectadas en membranas musculares parcialmente purificadas. Dos isoformas específicas han sido identificadas, clonadas y se ha encontrado que se expresan en el músculo esquelético. La isoforma GLUT 1, que se encuentra en poca cantidad en el músculo esquelético y otros tejidos, mientras que la isoforma GLUT 4 se expresa altamente en el músculo esquelético (quizás como un 95 % del total de transportadores), y se cree que es responsable de muchas propiedades que regulan el transporte de glucosa en las células.
Aunque las dos isoformas de transportadores comparten un 67 % de homología en términos de secuencia de aminoácidos, parecen tener propiedades reguladoras muy diferentes. Varios estudios han encontrado que los mecanismos que posee el músculo esquelético para incrementar el transporte de glucosa después del ejercicio, se producen a través de la migración de transportadores de glucosa GLUT 4, desde un sitio de almacenamiento intracelular hasta la superficie de la membrana celular. La evidencia morfológica directa usando anticuerpos y microscopio electrónico ha indicado que la isoforma del transportador GLUT 1 está presente a lo largo de la superficie de la membrana del músculo esquelético y que participa en el transporte basal de glucosa en ausencia de la insulina o de la contracción muscular. De otro modo, el transportador GLUT 4 está localizado en cercana proximidad al sistema de túmulos (T) transversales y puede migrar a la membrana del túbulo-T en respuesta a la insulina y quizá también a las contracciones musculares (3). Aunque los GLUT 4 se translocan a la superficie de la membrana como respuesta a la insulina, el contacto con la membrana de los túbulos-T puede tener consecuencias importantes durante el ejercicio. El sistema de túbulos-T provee un canal especial donde los sustratos transportados a la fibra muscular, pueden difundir dentro del centro del músculo sin vencer grandes distancias de difusión. El lúmen de los túmulos-T, es considerado suficientemente grande para la entrada de proteínas plasmáticas, además el transporte de sustratos extracelulares a través del sistema luminal, puede ser facilitado por las contracciones/relajaciones musculares, que pueden aumentar el flujo a través del extenso canal de túbulos. La presencia de receptores de insulina en la membrana de los túmulos-T, puede también ser un factor importante, que permite la entrada de glucosa dentro del músculo luego de la ingesta de carbohidratos.
ROL DE LA SENSIBILIDAD INCREMENTADA DE LA INSULINA EN LA RESÍNTESIS DE GLUCÓGENO MUSCULAR
El mecanismo por el cual el ejercicio y/o la insulina estimulan la translocación de transportadores de glucosa, es crítico para la regulación de la entrada de glucosa y por esto para la resíntesis de glucógeno post-ejercicio. El ejercicio está asociado a un cambio en la respuesta a la dosis de insulina, ya que una concentración reducida de insulina es requerida para obtener la misma velocidad de transporte de glucosa en comparación con un músculo no ejercitado. In-vivo, un solo período de ejercicio puede incrementar la sensibilidad a la insulina, siempre que los niveles de glucógeno sean disminuidos durante el ejercicio. Varios estudios han demostrado que el incremento en la sensibilidad a la insulina en todo el cuerpo en animales, puede ser ampliamente explicado por un incremento en los transportadores de glucosa presentes en la superficie de la membrana muscular después del ejercicio. En algunos casos, la insulina puede también incrementar la actividad intrínseca de transportadores de glucosa de la superficie, conduciendo a una actividad incrementada de los transportadores de glucosa. La sensibilidad a la insulina incrementada luego del ejercicio, juega un rol clave en la estimulación del transporte de glucosa y en el incremento de la actividad de la enzima glucógeno sintetasa en el músculo esquelético.
Aunque el ejercicio solo, estimula los dos procesos, la combinación de la insulina y el ejercicio resultan frecuentemente en un estímulo adicional al transporte de glucosa. Ha sido sugerido que existen dos fuentes de transportadores de glucosa, un grupo estimulado por el ejercicio y el otro por la insulina. Muchos estudios han sugerido que la sensibilidad incrementada a la insulina puede estar de algún modo acoplada a la depleción de glucógeno, ya que los efectos del ejercicio sobre la sensibilidad a la insulina son inversos a los de la repleción de glucógeno.
Los investigadores también han demostrado que el contenido de proteínas transportadoras de glucosa GLUT 4, es más de 5 veces más grande en los músculos rojos oxidativos, que en los músculos esqueléticos blancos. Esto no es sorprendente dado la gran diferencia en la velocidad máxima de transporte de glucosa estimulada por la insulina, en músculos esqueléticos rojos oxidativos, que puede ser causado en parte por una transcripción incrementada del gen GLUT 4. Esta adaptación, junto con un incremento similar de la actividad de la enzima glucógeno sintetasa I, puede ayudar a explicar la mayor velocidad de resíntesis de glucógeno observada en los músculos entrenados.
Un caso especial en el cual la resíntesis de glucógeno post-ejercicio es perjudicada, comprende al ejercicio excéntrico. El mismo, en cual el músculo se estira mientras desarrolla tensión, es un componente normal del ejercicio, pero resulta frecuentemente en un daño muscular y en dolor muscular post-ejercicio, en individuos desentrenados. De manera contraria al incremento de la sensibilidad a la insulina en todo el cuerpo, usualmente encontrada después del ejercicio. Ha sido encontrado que el ejercicio excéntrico está asociado a una sensibilidad a la insulina perjudicada, y a un fallo para resintetizar completamente el glucógeno muscular. Aunque el mecanismo responsable de la resíntesis de glucógeno perjudicada, es desconocido, la evidencia ultraestructural sugiere que el ejercicio excéntrico resulta en un daño en la superficie de la membrana muscular, que probablemente interfiere con el transporte de glucosa dentro del músculo esquelético y con la resíntesis de glucógeno. Esta explicación parece la más razonable, ya que el daño muscular no parece interferir con la actividad de la glucógeno sintetasa.
SEÑALIZACIÓN DE LA INSULINA Y DEPOSICIÓN DEL GLUCÓGENO
Los eventos moleculares que conectan la interacción de la insulina y su receptor, con la activación de los procesos sensibles a la insulina en las células, como el trasporte de glucosa y la activación de la síntesis de glucógeno, tienen que ser todavía claramente definidos. La evidencia reciente sugiere que la activación de la proteín serina/treonina quinasa, por la insulina, resulta en la activación de la fosfatasa I y su asociación con proteínas como la glucógeno sintetasa. Sin embargo, la conexión entre la activación del receptor de la insulina y la activación de estas proteín serina/treonina quinazas, es todavía confuso.
Varias proteín quinazas sensibles a la insulina parecen ser sustratos para los receptores de insulina, los cuales están regulados por las mismas, por medio de la fosforilación de la serina/treonina. Esto puede ser demostrado usando el inhibidor de la serina/treonina fosfato fosfatasa, ácido akadaico, que imita las acciones de la insulina, de incrementar la actividad de las proteín quinazas, y por ello estimulando la actividad de transporte de glucosa en el músculo esquelético. La contracción muscular también parece alterar la ruta de señalización de la insulina a través de la fosforilación de la proteína citoplasmática de 165-185 kd, llamada receptor de insulina sustrato-1 (IRS-1). La proteína IRS-1 tiene múltiples sitios de fosforilación, y ha sido demostrado, que es fosforilada por la estimulación de la insulina. Sin embargo, su rol para mediar la respuesta al ejercicio/contracción, tiene que ser todavía determinada.
Así, la principal señal responsable de la captación y el metabolismo de la glucosa incrementados después del ejercicio, así como la influencia de otros factores distintos a la insulina, necesitan todavía ser considerados. Estas preguntas permanecerán sin respuesta hasta que pueda ser sugerido un mecanismo celular detallado, que explique la habilidad del músculo esquelético de resintetizar glucógeno después del ejercicio.
Finalmente, los mecanismos responsables del fenómeno de supercompensación de glucógeno muscular después del ejercicio, todavía tienen que ser completamente explicados. Los requerimientos más importantes parecen ser la depleción de la reserva de glucógeno, la recuperación con una dieta alta en carbohidratos, y una respuesta normal de la insulina. Durante la supercompensación de glucógeno, al activación de la glucógeno sintetasa, podría teóricamente limitar la síntesis de glucógeno muscular. Sin embargo, la evidencia sugiere que el índice de actividad de la glucógeno sintetiza (A a I) es elevado solo durante el período, cuando los niveles de glucógeno retornan a la normalidad (24-36 horas) y no pueden explicar la supercompensación encontrada 48 horas después del ejercicio. Más probablemente la habilidad de estimular la supercompensación de glucógeno puede ser explicada en parte por la sensibilidad incrementada a la glucosa-6-fosfato, que activa la glucógeno sintetiza en la forma D (ver sección previa). Bajo estas circunstancias, la formación de glucógeno in-vivo es más probablemente conducida por un incremento de los transportadores de glucosa estimulados por la insulina. Tan pronto como la glucosa entra en la célula, es fosforilada a glucosa-6-fosfato por medio de la enzima hexoquinasa y debe ser utilizada a través de la glucólisis o almacenada como glucógeno. La deposición incrementada de glucógeno requiere que la glucosa no entre en la ruta glucolítica o experimente la oxidación hasta CO2 y H2O. La sensibilidad incrementada de la glucógeno sintetasa a bajos niveles de glucosa-6-fosfato puede de este modo permitir la activación de la enzima. Sin embargo, continúa por ser determinado si esta activación comprende niveles reducidos de la actividad de la fosfatasa, redisposiciones de la molécula de glucógeno, o algún otro mecanismo. Así, las bases bioquímicas para la resíntesis sobrenatural de glucógeno después del ejercicio prolongado, probablemente involucre una compleja interacción entre los carbohidratos de la dieta, el transporte de glucosa y la activación de la glucógeno sintetasa por medio de la insulina.
Agradecimientos: El apoyo para la preparación de este artículo fue otorgado por una Concesión de Desarrollo Profesional de la Juvenil Diabetes Foundation.
Dirección para correspondencia: Case Western Reserve University Department of Biochemistry 10900 Euclid Avenue Cleveland, Ohio 44106-4935 ph. (216) 368-6166 fx. (216) 368-4544
REFERENCIAS
1. Costill, D. L., W. M., Sherman, W. J., Fink, C., Maresh, and M., Witten (1981). The role of dietary carbohydrates in muscle glycogen resynthesis after strenuous running. Am. J. Clin. Nut; 34: 1831-1836
2. Johnson, J. L., and G. J., Bagby (1988). Gluconeogenic pathway in liver and muscle glycogen synthesis after exercise. J. Appl. Physiol;. 64:1591-1599
3. Friedman, J. E., R.W., Dudek, D., Whitehead, D., Downes, W., Frisell, J. F., Caro, and G. L. Dohm (1991). Immunolocalization of glucose transporter GLUT4 in human skeletal muscle. Diabetes; 40:150-154
4. Conlee, R.K., R. C., Hickson, W. W., Winder, J. M., Hagberg, and J. O., Holloszy (1978). Regulation of glycogen resynthesis in muscles of rats following exercise. Am. J. Physiol;. 235:R145-R150
5. Blom, P. C. S., N. K., Vollestad, and D. L., Costill (1986). Factors affecting changes in muscle glycogen concentration during and after prolonged exercise. Acta Physiol. Scand; 128:67-74
6. Ivy, J.L (1987). The insulin-like effect of muscle contraction. In Pandolf (Ed.) Exercise and Sports Sciences Reviews, Vol 15. pp. 29-51, MacMillan, New York
7. Kern, M. J. A., Wells, J. M, Stephens, C. W., Elton, and J. E., Friedman, E. B., Tapscott, P. H., Pekala, and G. L., Dohm (1990). Insulin responsiveness in skeletal muscle is determined by glucose transporter (GLUT4) protein level. Biochemical Journal; 270:397-400
8. Bak, J. F., and O., Pedersen (1990). Exercise-enhanced activation of glycogen synthase in human skeletal muscle. Am. J. Physiol;. 258:E957-963
9. Goodyear, L. J., M. F., Hirshman, P. A., King, E. D., Horton, C. M., Thompson, and E. S., Horton (1990). Skeletal muscle plasma membrane glucose transport and glucose transporters after exercise. J. Appl. Physiol; 68:193-198
10. Cartee, G. D., D. A., Young, M. D., Sleeper, J., Zierath, H., Wallberg-Henrikkson, and J. O., Holloszy (1990). Prolonged increase in insulin-stimulated glucose transport in muscle after exercise. Am. J. Physiol; 256:E494-E499
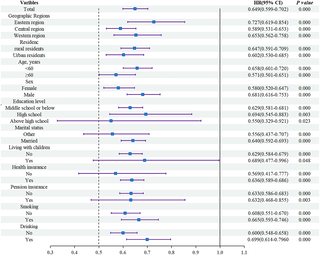
Asociación entre trayectorias de capacidad intrínseca y riesgo de incidencia de accidente cerebrovascular en adultos chinos de mediana edad y mayores: Evidencia de un estudio de cohorte prospectivo a nivel nacional basado en CHARLS
La Influencia de la Capacidad Intrínseca en el Riesgo de Accidentes Cerebrovasculares en Adultos...
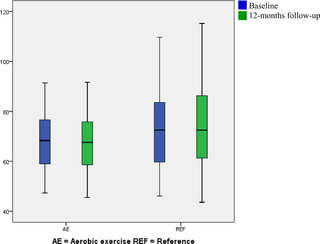
Efectos a Largo Plazo sobre los Factores de Riesgo para Enfermedades Cardiovasculares después de 12 Meses de Intervención de Ejercicio Aeróbico – Un ECA en el Lugar de Trabajo entre Limpadores
La investigación sobre la salud cardiovascular de limpiadores ha adquirido relevancia, dado que...
La evidencia emergente y en evolución que respalda la creatina como ayuda ergogénica: historia y aplicaciones
Resumen Antecedentes: La creatina es una de las ayudas ergogénicas más estudiadas, con más de tres...
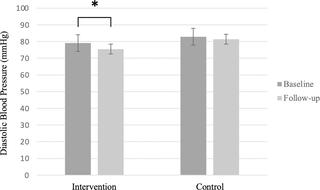
Efecto del ejercicio isométrico de agarre manual en casa con un dispositivo comercial en la presión arterial en adultos mayores con hipertensión: un ensayo controlado aleatorio
Efecto del Ejercicio Isométrico de Mano en Casa sobre la Presión Arterial en Ancianos con...
El ejercicio aeróbico atenuó la fibrosis pulmonar inducida por bleomicina en ratones dominantes en Th2
El ejercicio aeróbico (AE) ha ganado atención por su potencial terapéutico en trastornos...